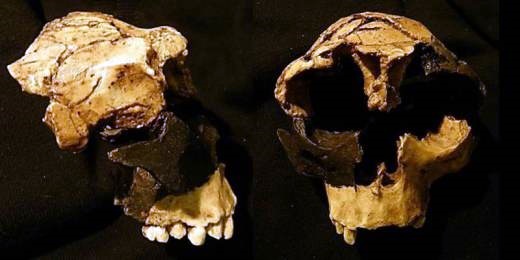
Homo antecessor. Composición de ATD6-15 y ATD6-69. Fotografía de Roberto Sáez
Hace casi veinticinco años que el equipo de Atapuerca, liderado por los paleoantropólogos Juan Luis Arsuaga, Jose María Bermúdez de Castro y Eudald Carbonell, anunciaba oficialmente el descubrimiento de una nueva especie en el genero Homo a la que denominaron ‘antecessor’, ‘pionero’, término que Bermúdez de Castro escogió al rememorar las avanzadillas que el Imperio Romano enviaba a tierras ignotas. La noticia revolucionó a la comunidad científica y puso al yacimiento burgalés en la primera línea de la investigación paleoantropológica mundial. Sin embargo, la juventud del espécimen ATD6-69, base de sus investigaciones, pronto llevaría a otros colegas de profesión a plantearse serias dudas acerca de la viabilidad de antecessor como nueva especie. Y surgió la pregunta, ¿existió realmente Homo antecessor?
Profesor Lee Berger, con una réplica de cráneo naledi
Homo naledi se encuentra entre los hallazgos arqueológicos más recientes relacionados con la Prehistoria, entre aquéllos que, posiblemente, nos obliguen a reconfigurar nuestro árbol genealógico y a reescribir el itinere de nuestra propia evolución.
La Prehistoria se encuentra en constante revisión. El hallazgo de nuevos yacimientos, con la consiguiente aparición de otros tantos fósiles, y lo refinado de las últimas técnicas de análisis, cada vez más innovadoras, permiten acotar con más y más precisión unas cronologías que parecen retroceder en el tiempo sin encontrar su límite inferior. Sin embargo, a tenor de las opiniones de los protagonistas de esta historia, el caso naledi plantea una dicotomía cronológica que hace dudar a estos mismos especialistas.
Profesor John Hawks
Pero no es ése el gran desafío, pese a la importancia de la cronología, no lo es. Es el de intentar explicar aquellas manifestaciones que no se corresponden con tecnocomplejos, que no suelen dejar un rastro material, y que tienen mucho que ver con pensamientos elevados y con estados de conciencia fuera de lo habitual, caso de las prácticas rituales, básicamente funerarias.
Como veremos, a continuación, en el artículo que os hemos traducido, Homo naledi aglutina todas esas facetas, aquéllas que nos distinguen del resto de la Creación, aquéllas que, en definitiva, nos hacen humanos, y que sin embargo, no podrían corresponderse con un cerebro tan diminuto, al menos, en teoría.
¿Será Homo naledi el eslabón perdido, o su clave misma? Con el tiempo lo descubriremos.
De: Paul Rincon, Responsable de Ciencias de BBC News, 25 April 2017
Traducción: LPDLA
Un tipo de humano primitivo, del que una vez pensamos tendría una antigüedad de tres millones de años, vivió en realidad en época mucho mas reciente, tal y como sugiere un estudio.
Los restos de quince esqueletos parciales pertenecientes a la especie Homo naledi fueron descritos en 2015, tras ser hallados en un sistema de cuevas en Sudáfrica por un equipo dirigido por Lee Berger, procedente de la universidad de Wits, quien, en una entrevista, sostiene ahora que la antigüedad de los mismos se estima entre los doscientos mil y los trescientos mil años.
Aunque su anatomía comparte ciertas semejanzas con la de los humanos modernos, otras características de Homo naledi nos sugieren que aquellos humanos vivieron en épocas mucho más tempranas, hace unos dos millones de años, o incluso más.
“Son como una forma primitiva de nuestro propio género, el género Homo; al parecer, podrían compartir linaje con los primeros Homo erectus u Homo habilis, o con Homo rudolphensis“, ha comentado John Hawks, de la Universidad de Wisconsin, colega del Profesor Berger.
Aunque algunos expertos consideraron que naledi pudo haber vivido en épocas relativamente recientes, el Profesor Berger declaró en 2015 a BBC News que los restos podrían alcanzar los tres millones de años. Nuevas pruebas de datación sitúan a la especie en una cronología en la que Homo naledi pudo haber coincidido con los primeros ejemplares de nuestra especie, Homo sapiens.
En el programa de radio de la BBC, Inside Science, el Profesor Hawks ha afirmado que “presentan la edad de los Neandertales en Europa, la de los Denisovanos en Asia, la de los primeros humanos en África. Son parte de esta diversidad mundial que ya estaba presente, al tiempo que nuestra especie surgía”.
“Ignoramos que más hay en África que podamos encontrar, ese es el gran mensaje, en mi opinión. Si este linaje, que al parecer se originó hace dos millones de años, estaba aún presente hace unos doscientos mil años, entonces es posible que no acabara ahí. No hemos encontrado al último naledi, hemos encontrado un naledi“.
Los restos de naledi fueron excavados en 2013 en el interior de una cámara de difícil acceso en el centro de un entramado de cuevas conocido como Rising Star. Por entonces, el profersor Berger se mostró convencido de que habían sido depositados en la cámara de forma deliberada, quizás durante generaciones.
Esta idea, que sugería que Homo naledi exhibía un comportamiento ritual, no estuvo exenta de controversia, pues tales prácticas son consideradas por algunos autores características de los humanos modernos.
El profesor Hawks ha dado a conocer que el equipo viene explorando una segunda cámara desde entonces. “La (segunda) cámara presenta los restos de, al menos, otros tres individuos, incluyendo un esqueleto parcial con su cráneo”.
Los investigadores ya han intentado extraer ADN de los restos a fin de conseguir más información sobre la posición ocupada por naledi en el árbol genealógico humano, si bien no han tenido éxito, por el momento.
“Los restos se hallan en unas condiciones que nos permiten considerar tal posibilidad. La cavidad es relativamente cálida si la comparamos con aquéllas del norte de Europa y Asia, donde la preservación del ADN es excelente”, ha señalado el profesor Hawks.
Se prevé la publicación en los próximos meses de un estudio concluyente sobre las pruebas de datación.
Paul Rincon
Si habéis llegado hasta aquí, qué mejor, creemos, que os quedéis un rato más con nosotros, para ver el documental “Dawn of Humanity”, “El Amanecer de la Humanidad” (2015), donde se muestra cómo se llevó a cabo todo el trabajo previo de preparación de las excavaciones, y cuán laborioso resultó el proceso de recuperación de los restos, tarea llevada a cabo por un fenomenal equipo femenino.
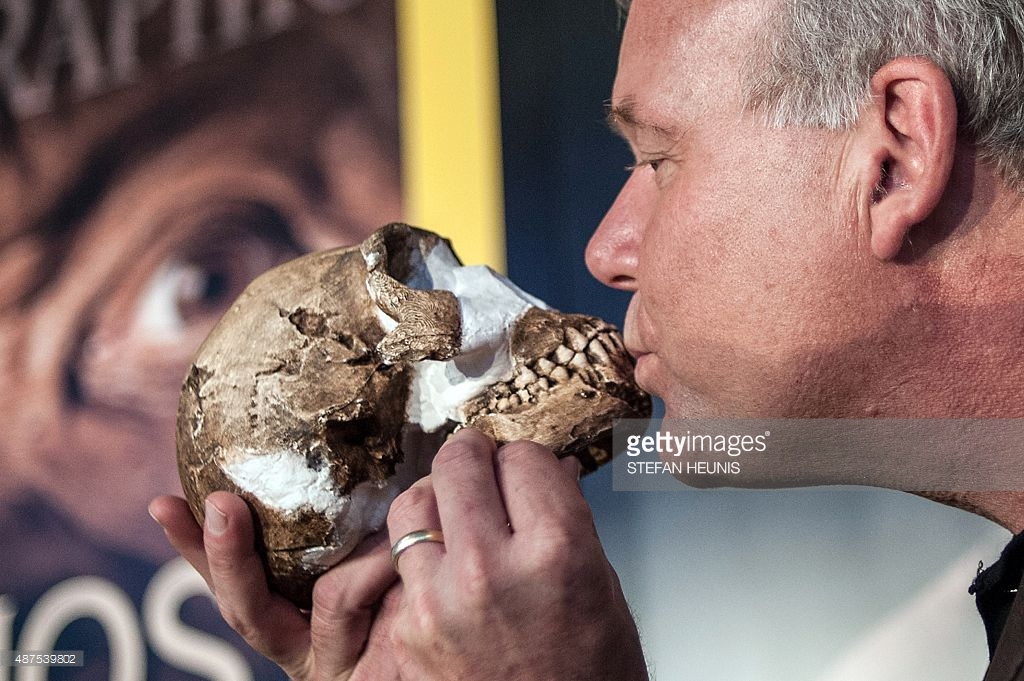
Berger besa una réplica de un cráneo naledi, un ancestro humano recién hallado durante los descubrimientos que se desvelaron el 10 de Septiembre de 2015 en Maropeng, Sudáfrica. Una nueva especie, Homo naledi, con una antigüedad entre 2,5 y 2,8 millones de años, fue descubierta en la denominada “Cuna de la Humanidad”, a unos cincuenta kilómetros al Noroeste de Johannesburgo. De unos cinco pies de altura y unas cien libras de peso, aproximadamente, y con un cerebro no mayor que el tamaño de media naranja, Homo naledi es una combinación sorprendente de características propias, tanto de australopitecos como de humanos, que era absolutamente desconocida para la Ciencia, hasta ahora, como afirman los investigadores