“El mar dará a cada hombre una nueva esperanza, como el dormir le da sueños”
Cristoforo Colombo, Cristóbal Colón
Contexto histórico
El Doce de Octubre de 1492, tras una larga y penosa travesía por el Océano Atlántico, las tres carabelas comandadas por el almirante y cartógrafo genovés fondeaban en la isla de Guanahani, que pertenece al archipiélago de las Bahamas y que, en la actualidad, conocemos por San Salvador.
Creyendo haber llegado a las Indias, la hazaña de Colón y de los tripulantes de la Santa María, La Pinta y La Niña marcaría el inicio de la Edad Moderna, la Época de los Descubrimientos, y sentaría las bases de un Imperio, el español, donde el Sol nunca se ponía. Pero implicó mucho más: el Descubrimiento de América devino en lo que conocemos por Hispanidad, una auténtica globalización, tal y como muchos expertos consideran. Un inmenso legado que permanecerá siempre, pese a los intentos de ciertos lobbies de defenestrarlo.
Cinco siglos después, cuando hablamos de la conquista espacial, los que en ella están inmersos se refieren al Descubrimiento de América para intentar explicar sus motivaciones. En ese sentido, el espíritu de aventura, de alcanzar nuevas fronteras, de llevar a la Humanidad un paso más allá, no ha cambiado, sigue siendo el mismo, sea atravesando el frío y vasto océano o las profundidades del espacio exterior.
Cuentan las crónicas que esta fue la arenga con la que el General y estadista cartaginés Aníbal Barca (247 – 183 a.C.) lanzó a sus tropas a la Segunda Guerra Púnica contra Roma en 218 a.C. Las mismas que un año antes le habían dado la victoria contra Sagunto, permitiéndole el control de la costa levantina.
Perfectamente pertrechados con manteletes – grandes escudos de madera ensamblados como un rompecabezas y de formas diversas, cuadrangulares, circulares -, ballestas, y picos, los soldados cartagineses, con Aníbal al frente, derribaron las murallas de la ciudad mediante el empleo de catapultas, parapetos móviles y las torres de asalto que los griegos llamaban helepolis, las tomadoras de ciudades.
Se iniciaba, así, un nuevo conflicto bélico entre Cartago y Roma, que Aníbal prepararía desde su residencia en Qart Hadasht encomendándose al dios Melqart.
A finales del año pasado, Taiwán alertaba a la Organización Mundial de la Salud del brote en la ciudad de Wuhan, en la China continental, de una extraña afección respiratoria que cursaba con cuadros de neumonía y que era altamente contagiosa. No sería hasta el día 31 de diciembre cuando su director general, el etíope Tedros Adhanom, a quien sus detractores acusan de haber ocultado dos epidemias de cólera en su propio país y de una actitud indulgente en exceso para con un régimen tan poco transparente como el chino, reconociese públicamente la gravedad de la situación. Una decisión tardía que los enormes intereses comerciales que China tiene en todo África y en Etiopía, especialmente, por sus ricos yacimientos en tierras raras, materia prima fundamental para que el gigante asiático pueda proseguir con la fabricación de dispositivos móviles a gran escala, podrían explicar.
Semana tras semana, hemos visto cómo el brote se ha expandido por los cinco continentes, habiendo provocado cientos de miles de fallecidos, de contagiados, de pacientes con secuelas, y el colapso sanitario en los países que peor han gestionado la situación. Un escenario que nos ha hecho recordar aquella epidemia de la mal llamada Gripe Española y que en lo económico nos sitúa en una recesión peor que la Gran Depresión de 1929. El impacto, del que todavía desconocemos sus últimas consecuencias, ha sido brutal y ha conseguido que el mundo se paralizara en estos primeros meses de 2020.
Mercado de Huanan I
Como suele ocurrir con ciertas catástrofes, se ha especulado mucho sobre si el origen del patógeno era natural o artificial. Unos piensan que la pandemia ha sido consecuencia de una zoonosis, un contagio entre especies animales que, en última instancia, ha alcanzado al ser humano. El foco del brote, según esta hipótesis, estaría localizado en el mercado de abastos de Huanan, en la ciudad de Wuhan, en el que numerosas especies animales son vendidas, aún vivas, para su consumo. El de Huanan es uno de tantos de los denominados mercados húmedos, es decir, establecimientos sin control sanitario alguno, sin un mínimo de condiciones higiénicas, en donde los animales ya sacrificados son exhibidos junto a los ejemplares vivos que esperan la muerte en jaulas insalubres. La oferta es de lo más variada: civetas, zorros, salamandras, pavos reales, ratas, cachorros de lobo, koalas, cocodrilos, puercoespines, erizos, venados, tejones, murciélagos y pangolines.
Mercado de Huanan II
Otros consideran, por contra, que el virus se ha escapado del laboratorio de experimentación biológica de nivel 4 de seguridad que, curiosamente, queda muy cerca de ese mercado y donde se viene realizando toda suerte de ensayos con diferentes tipos de coronavirus. Desde este espacio no tenemos la capacitación necesaria para afirmar la veracidad de una u otra explicación, y por ello pensamos que lo mejor es remitirse a los expertos de verdad.
Mercado de Huanan III
Entrevistado en el canal de Youtube Milenio Live, de los periodistas Iker Jiménez y Carmen Porter, quienes han arrojado luz como nadie en los medios de información españoles gracias a contar con los científicos y especialistas más prestigiosos, el Dr. Adolfo García – Sastre, virólogo y Jefe de Patología del Hospital Monte Sinaí, en Nueva York, afirmó que los coronavirus tienen un origen natural y que los murciélagos, entre otros animales, son posibles vectores del contagio. Preguntado por la posiblidad de que el covid – 19, acrónimo de coronavirus disease 2019, como ha sido bautizado, hubiera escapado de ese laboratorio, el buen doctor respondió que no se podía descartar de forma tajante, pero, que “el virus ya estaba ahí fuera”. De sus palabras, deducimos que, casi con toda probabilidad, nos encontramos ante una zoonosis.
Covid – 19, no es el único estallido viral que se ha producido en años recientes. Recordemos otros como los de SARS, acrónimo de Severe Acute Respiratory Syndrome, en 2003, y de MERS, acrónimo de Middle East Respiratory Syndrome, oriundos de las selvas asiáticas y de Oriente Medio, respectivamente. Pero el objeto de este artículo no es entrar en disquisiciones científicas que para nada dominamos, sino aproximarnos a los antecedentes históricos de unos brotes que son consecuencia de ciertas prácticas de consumo y culinarias cuyas razones explicaremos a continuación.
El Gran Timonel
«Cuando no hay comida suficiente, la gente muere de hambre. Merece la pena que la mitad muera para que la otra mitad pueda comer bien»
Mao Tse – tung
El descontento provocado por la situación en que China había quedado frente a Japón tras la paz de 1918, que aquélla se negó a firmar, provocó la aparición en 1919 del Movimiento del Cuatro de Mayo. Lo que empezó como unas manifestaciones estudiantiles, que se saldaron con la dimisión de rectores universitarios, se transformó en una movilización que acabaría por extenderse por todos los rincones del país. Un movimiento en el que un joven ayudante bibliotecario de la Universidad de Pekín, Mao Zedong, más conocido como Mao Tse – tung, participaría activamente. En 1920, las revistas universitarias ya empezaban a incluir en sus páginas textos de índole marxista, al tiempo que la primera traducción al chino de El Manifiesto Comunista era editada.
Pero el marxismo, lejos de proporcionar unidad, iba a agudizar la división entre los sectores reformistas de la política china. En 1921, se fundaba en Shanghai el Partido Comunista Chino con el beneplácito de diferentes delegados del país cuyos nombres no se conoce con exactitud, salvo el de Mao Tse – tung. El proyecto político de Mao y de sus seguidores pasaba por atraer a las masas rurales al comunismo. En un país donde la obediencia a la clase dirigente forma parte de su acervo cultural, la vanidad y las visiones megalómanas de Mao le reforzaron en el poder hasta tal punto que, hacia 1935, era ya el indiscutible líder supremo del partido y le apodaban el Gran Timonel.
El sufrimiento de un pueblo
En 1955, millones de campesinos fueron desplazados a la fuerza hacia comunas en un proceso de colectivización orientado hacia la eliminación absoluta de la propiedad privada. Un plan tan plagado de incompetencias como de medidas lesivas sólo podía tener consecuencias catastróficas, de manera que la producción se desplomó. Pero lo peor estaba aún por llegar.
En 1958, con el denominado Gran Salto Adelante, la presión sobre las comunas fue en aumento. La de Mao era una iniciativa fracasada desde el mismo momento de su concepción, que sumiría en el hambre y la miseria a vastas regiones del país y provocaría la muerte de unos cuarenta millones de personas, según los últimos estudios. En su búsqueda de los culpables, los sufridos agricultores y los funcionarios del partido no fueron los únicos en ser puestos en el punto de mira del líder comunista.
Guerra a los gorriones I
Mao achacó a los gorriones el descenso drástico en la producción de grano y ordenó su exterminio con todos los medios posibles. Estos pajarillos, que no sólo se alimentan de cereales, sino de insectos, también, y realizan una labor maravillosa como plaguicida natural, fueron combatidos de diversas maneras que pasaban por abatirlos con munición, por espantarlos para impedirles posarse, manteniéndolos en vuelo para matarlos por puro agotamiento y por envenenamiento, a base de cebos que terminarían por contaminar la tierra y matar a otras muchas especies.
Guerra a los gorriones II
La cacería de gorriones llegó a tal extremo que los biotopos perdieron su equilibrio natural. Las plagas, especialmente de langosta, se descontrolaron y arrasaron las plantaciones. Cuando las autoridades repararon en su tremendo error, la población de estas aves estaba tan al límite de su extinción que tuvieron que importar en secreto ejemplares de la U.R.S.S. Recurrieron al uso de plaguicidas en tal cantidad que las cadenas tróficas quedaron afectadas. Se produjeron inundaciones tras la que vinieron sequías que agudizaron el hambre de una población tan desesperada que se vería abocada incluso al canibalismo.
La persecución a los trabajadores comunales y a los miembros del partido fue implacable. Los primeros sufrieron maltratos continuados por los cuales unos dos millones y medio de campesinos perecieron. Entre los segundos, los considerados responsables fueron depurados. Sin embargo, las evidencias de lo que realmente había sucedido fueron recopiladas por altos cargos dentro del partido y la posición de Mao se resintió.
Hornos de acero en las comunas
El Gran Salto Adelante, entre 1958 y 1962, se tradujo en un “descenso a los infiernos”, según el historiador holandés, especializado en la Historia de China, Frank Dikötter en La Gran Hambruna en la China de Mao. Una guerra sin cuartel a la Naturaleza, desviando el curso de ríos, derrumbando montañas y poniendo a especies al borde de su extinción, caso de los gorriones. A nivel social, se impuso el culto a la personalidad, la fe ciega en el líder y se laminó todo atisbo de disidencia. En lo económico, el intento de industrialización del país a marchas forzadas, empezando por el propio sector agrario, igualmente fracasó.
Revolución Cultural I
Con su primera prueba nuclear en 1964, China entraba en el exclusivo club de las potencias armamentísticas. La demografía seguía en aumento pese a que el hambre continuaba haciendo estragos entre una población cada vez más mermada. Por si todo esto no hubiese sido suficiente, la bautizada como Revolución Cultural (1966 – 1969) terminó por fracturar al país. Cárcel, torturas, trabajos forzados y condenas a muerte se convirtieron en el pan de cada día. Los intelectuales fueron masivamente depurados y las universidades clausuradas. Nadie, absolutamente nadie, con independencia de su profesión o de su estamento social, estaba libre de ser perseguido por la temida Guardia Roja. Ni siquiera una de las instituciones clave en la sociedad china como era la familia quedaría a salvo de los ataques del partido en aras de la “modernización” que propugnaban.
Revolución Cultural II
Mao Tse – tung falleció en 1976. Fue sucedido por Hua Guofeng quien acabaría cediendo el poder a Deng Xiaoping, un dirigente de corte pragmático que renunciaría a las políticas revolucionarias al tiempo que reforzaría el modelo centralista y autoritario del Estado. Deng propulsó las reformas necesarias que han permitido, desde entonces, un intenso crecimiento económico que ha colocado al país como ejemplo de un modelo de capitalismo “salvaje” controlado por una élite comunista. La misma que sigue impidiendo un sistema constitucional y democrático de partidos, la celebración de elecciones libres y movimientos de oposición al partido único.
Si habéis llegado hasta aquí, estaréis preguntándoos qué tiene que ver este episodio de la Historia Contemporánea de China con la pandemia provocada por el coronavirus.
Mao, Stalin, Lenin, Engels, Marx
Cuando Mao se dio cuenta del fracaso económico que el Gran Salto Adelante había supuesto, de las hambrunas que había provocado y de la pérdida de su prestigio personal, que estaba ya muy cuestionado, permitió a las granjas comunales la cría y venta de todo tipo de especies de animales salvajes, como las que hemos mencionado al principio de este artículo. Hacinándolos de forma insalubre y procesando su carne sin las mínimas garantías sanitarias, estos campesinos facilitaron de manera inconsciente la transmisión de enfermedades entre especies. Como las desgracias nunca vienen solas, el pueblo chino no conocía lo que era disponer de un simple frigorífico en casa hasta hace muy pocas décadas, razón que explica su apetencia por el consumo de animales vivos.
Con la llegada de Deng Xiaoping al poder en 1978, esa economía paralela, lejos de ser abolida, continuó. Lo que había empezado como una práctica de subsistencia en las comunas, se afianzó en las ciudades a través de grandes mercados en los que los animales eran sacrificados en el momento de su compra por parte del consumidor en unas condiciones deplorables. Una situación explosiva cuya primera detonación se produjo en 2003, cuando el brote de SARS se extendió por más de setenta países y acabó con la vida de más de setecientas personas, cifras tan lamentables como inferiores a las provocadas por este coronavirus. En un primer momento, el Gobierno chino consideró la prohibición de estas actividades, pero cedió ante las presiones de los sectores implicados. Lejos de corregir el error, la lista de especies fue ampliada en 2016 para incluir al tigre y al famoso pangolín.
Deng Xiaoping
Desconocemos que papel desempeñan, si es que tienen alguno, tanto la Organización Mundial de la Salud como Naciones Unidas en todo este asunto, cuya responsabilidad es tan grande como su impericia. Que las ideas perniciosas de una mente calenturienta y megalómana como la de Mao Tse – tung hayan tenido unas consecuencias sanitarias, sociales y económicas tan dramáticas como las que estamos viviendo casi medio siglo después, es muestra de los peligros del totalitarismo y de una globalización mal concebida desde sus inicios, que nos ha hecho tan dependientes de un país que ha de progresar mucho aún en cuanto a transparencia, derechos y libertades y sanidad. Incluso con la llegada de la ansiada vacuna o de, al menos, un tratamiento paliativo como los que empleamos para la gripe común, el peligro no habrá pasado del todo, a menos que los acuerdos internacionales consigan que el Gobierno chino de por prohibidas esas prácticas de consumo que siguen poniendo en riesgo a sus sufridos ciudadanos y al conjunto de la población mundial.
Flamen era el sacerdote romano encargado del culto a una deidad determinada. El flamen Augustal dirigía el culto a Augusto, primer emperador romano, divinizado por el Senado tras su muerte en el año 14 d.C. Del culto a Júpiter, dios de la Luz, mediador entre dioses y hombres, se responsabiliza el flamen Dial. El flamen Marcial hacía lo propio respecto del culto a Marte, dios de la Guerra. El flamen Quirinal era responsable del culto a Quirino, como se conoció a Rómulo, fundador de Roma, tras ser divinizado, si bien no todos los historiadores consideran que se trata del mismo personaje.
Su vestimenta consistía en un largo manto abrochado al cuello y un gorro a modo de casquete rematado por un borlón de lana. En sus manos, un bastón fabricado en madera de olivo.
Las esposas de los flámines eran las flamínicas. Consideradas sacerdotisas, disfrutaban de numerosas prerrogativas y exhibían avalorios distintivos.
En 1989, el común de los mortales no teníamos ni la más remota idea de qué era Internet. El término era desconocido para la mayoría de nosotros, con la excepción, quizás, de aquellos aficionados a MS Dos – ese sistema operativo tan incipiente a cuyo lado el Pleistoceno nos parece moderno – y a los famosos ordenadores Spectrum, unos primeros usuarios informáticos a quienes muchos veían como bichos raros que manejaban lenguajes extraños. Y es que habría de transcurrir toda una década para que el término se popularizara y los ordenadores irrumpieran en nuestras vidas de tal forma que, en el momento en que nos encontramos, no tener acceso a la red de redes desde un ordenador o desde un teléfono móvil nos resulta inconcebible. Tal ha sido su impacto que los expertos consideran que Internet representa la Tercera Revolución Industrial, y ya vislumbran la Cuarta. Esta reflexión viene a cuento del Día Mundial de Internet, que se celebra, tal como hoy, cada 17 de mayo.
Alan Turing
Como muchos otros grandes inventos que surgieron de la investigación militar y aeroespacial, Internet tiene sus orígenes en plena Guerra Fría entre la Organización del Tratado del Atlántico Norte (OTAN) y el Pacto de Varsovia, cuando Estados Unidos buscaba desarrollar un sistema que permitiera el acceso inmediato a la información desde cualquier ubicación en caso de un hipotético ataque soviético. Pero los primeros pasos de lo que por Internet conocemos son anteriores a esa época.
Al poco de iniciarse la Segunda Guerra Mundial, los servicios de inteligencia de las potencias en liza se plantearon la necesidad de desarrollar máquinas que, desde cálculos de probabilidad, descifraran mensajes codificados a velocidades que superaran con creces la capacidad del ser humano. El británico Alan Turing, creador de la máquina que lleva su nombre, conseguiría desencriptar Enigma, el sistema de comunicaciones alemanas, acortando la duración de la contienda y sentando de esta manera las bases de la Computación, fundamento de todo lo que vendría después. No sabremos nunca qué habría opinado el bueno de Groucho Marx si viviera hoy y alguien le recordara en un día como este aquella famosa cita suya en la que afirmó que “inteligencia militar son términos contradictorios”.
Tim Berners – Lee
Si hoy disfrutamos de todas las ventajas de Internet, es gracias a la sagacidad de Tim Berners – Lee, científico británico especializado en Computación y creador de la World Wide Web, con cuyo acrónimo www empiezan todas las páginas en Internet. En noviembre de 1989, Berners – Lee consiguió la primera comunicación entre un servidor y su cliente mediante un protocolo Hyper Text Transfer Protocol, o http. El camino hasta hoy es de sobra conocido. Internet ha supuesto una revolución en la forma en que nos comunicamos, compramos y conseguimos información. Ha dado voz a todo aquel que tenga algo que decir y ha posibilitado la irrupción de nuevos medios de comunicación tan alternativos como independientes que están poniendo en jaque a los considerados tradicionales. Para los creadores de contenido, la red se ha convertido en una herramienta de lo más útil para dar a conocer sus obras de manera exponencial, y eso que sólo un cincuenta y tres por ciento de la población mundial tiene acceso a Internet. Las denominadas redes sociales han sido determinantes en todo ese tráfico ingente de información.
Pero, aunque muchos usuarios no sean conscientes de ello, el espacio de comunicación y de libertad, sobre todo, que Internet representa, está muy amenazado. Como si de una encarnación del Gran Hermano de la novela de George Orwell “1984” se tratara, gobiernos y corporaciones diversas no escatiman esfuerzos en intentar poner puertas al campo. La pandemia sanitaria por coronavirus (covid – 19) que estamos sufriendo ha puesto en evidencia la labor censora y de desinformación que ciertas oficinas de verificación, vasallas del poder de turno, vienen ejerciendo cuando las publicaciones no son favorables a los señores a los que sirven. Y como David contra Goliat, en esa lucha está Tim Berners – Lee, una empresa en la que el padre del invento se ha embarcado con el propósito de devolver Internet a la ciudadanía a través de sus propuestas Contract for the Web, Solid e Inrupt, que se resumen en que el acceso a la libre información es un derecho y que éste no puede comprometer la privacidad de los usuarios. El tiempo nos dirá si lo consigue o no.
Por más que algunos se empeñen en hacer un uso maniqueo de la Historia, que no es otro que aquel que tiende a reducir la realidad, el papel de los colonizadores vascos y catalanes en la Conquista española de América fue determinante a la hora de hacerse con tan vasto territorio.
Las gestas españolas en América del Norte no son tan conocidas. Es por ello que os traemos hoy la historia de Gaspar de Portolá – quien impidió el avance de rusos y británicos en la Alta California – y de otros muchos compatriotas catalanes, gracias a una excelsa publicación de The Hispanic Council, un centro de estudios, o laboratorio de ideas, un think tank, que dicen los sajones, si bien nosotros siempre preferimos emplear nuestra propia lengua, que es más rica que la inglesa en muchos aspectos, y prescindir de tanto anglicismo.
The Hispanic Council se dedica, entre otras actividades, a la investigación y divulgación históricas a través del estudio de los lazos culturales que nos unen con EE.UU., un legado de más de cuatrocientos años de antigüedad desde que Ponce de León desembarcara en la costa de Florida en 1513 y cuyo esencia sigue viva a través de la toponimia, los símbolos y, por supuesto, la lengua.
Irenarca. Del latín tardío irenarcha, y éste del griego εἰρηνάρχης eirēnárchēs, de εἰρήνη eirḗnē ‘paz’ y -άρχης -árchēs ‘el que rige’.
Magistrados Romanos
Irenarca era el magistrado del Imperio romano encargado de velar por la paz y la tranquilidad del pueblo, sobre todo en las provincias romanas de Asia (formalizada a partir del año 133 a.C., cuando el rey de Pérgamo, Átalo III, lega su reino a Roma ante la falta de un sucesor al trono) y de África (que Roma conforma en el año 146 a. C. tras derrotar a Cartago en la Tercera Guerra Púnica y haciendo de Útica su capital), sobre todo en Egipto.
Así, el término irenarquía se refiere a la dignidad o jurisdicción de irenarca.
Gracias al programa Comando Actualidad, de La 1 de Radio Televisión Española, emitido en la noche de ayer bajo el título “Patrimonio en Peligro”, muchos televidentes tuvieron la oportunidad de descubrir una situación que, si bien nunca ha copado las portadas de los principales medios de comunicación nacionales, viene siendo reiteradamente denunciada por la Asociación Hispania Nostra, que se constituyó allá por 1976 con motivo del Año del Patrimonio Arquitectónico Europeo. Dicha situación les ha llevado a elaborar una Lista Roja de nuestro Patrimonio Histórico y Cultural.
A lo largo de la grabación, que podéis encontrar al final del artículo, el equipo del programa se traslada a una serie de localidades que albergan diferentes monumentos que comparten un denominador común, que no es otro que el ruinoso estado de sus estructuras. Si bien desconocemos con qué criterio se ha procedido a su selección, abarcar todos los enclaves habría sido imposible, dado que la lista contiene algo más de novecientos monumentos en serio peligro de desaparición.
Cierto es que la primera causa de este desaguisado se remonta al año 1836, cuando el ministro Juan Álvarez Mendizábal puso en marcha su famosa Desamortización, que permitió en su momento que muchos particulares se hicieran con elementos histórico patrimoniales que, hasta la fecha, habían pertenecido en su mayoría a la Iglesia. Personas, ellas y sus herederos, que con el paso del tiempo se desentenderían del mantenimiento de los mismos, bien por falta de interés, bien por escasez de medios económicos, o por ambas razones.
Lo interesante del asunto, y aquí viene nuestra reflexión, es que muchos de esos edificios históricos que parecen condenados al derrumbe se encuentran en la tan cacareada, últimamente, por parte de nuestros políticos, España Vacía o Vaciada. No vamos a entrar, ahora, en las razones por las cuales una comarca queda despoblada, quizá, en otro artículo lo hagamos, pero, si realmente sus señorías quisieran revitalizar esas localidades y ponerlas en el mapa de la sostenibilidad por la que tanto abogan, lo tendrían muy fácil: Bastaría con gestionar a través de las respectivas Comunidades Autónomas las ayudas económicas que permitieran volver a poner en valor toda esa parte de nuestra historia que se nos va.
Tal y como se describe en el reportaje, los importes necesarios para acometer los trabajos de rehabilitación y restauración pertinentes oscilan entre los treinta mil y el millón de euros. Cantidades que no pueden ser aportadas por municipios cuya media de población se sitúa en torno a los treinta habitantes, como mucho, pero que para las administraciones autonómicas y estatal son una nimiedad. El efecto sería doble, al recuperar unos edificios que, más pronto que tarde, sólo permanecerán en el recuerdo de unos pocos, y garantizar la supervivencia de unas poblaciones que podrían reinventarse gracias al Turismo Cultural, un sector que no ha dejado de crecer en los últimos años.
¿Recogerán sus señorías el guante a fin de que la mencionada lista torne en un color más esperanzador? Ese es el deseo de muchos españoles, nuestro, también. Sólo el tiempo lo dirá…
El pasado día 22 de Enero, el director de la Real Academia Española de la Lengua, Don Santiago Muñoz Machado, afirmó la impecabilidad gramatical de la Constitución Española ante la intención del Gobierno de proceder a “retoques estéticos” del texto mediante el denominado “desdoblamiento de género” y en aras a conseguir que también nuestra Carta Magna refleje ese uso “inclusivo” del lenguaje al que nos quieren someter. Una tendencia que, desde hace unos años, políticos, periodistas e incluso algunos docentes vienen recitando como si de un mantra religioso se tratase. Repite algo hasta la saciedad, que seguro que termina por calar, que diría aquel.
Tal es así que, cada vez con más frecuencia, tenemos que soportar aquello de “los españoles y las españolas”, “los trabajadores y las trabajadoras”, “los diputados y las diputadas”, en vez de “los españoles” – ellas y ellos -, “los trabajadores” – ellas y ellos -, “los diputados” – “ellas y ellos”. Pero, no nos extrañemos de que, en una época en la que la Educación está cada vez más manoseada y desprestigiada, en la que la gran cantidad de información a la que podemos acceder está provocando el efecto contrario, el de una ignorancia y un desconocimiento supinos, pensar que alguien llegue a considerar el Principio de Economía del Lenguaje, si es que acaso lo conoce, sea tan utópico como improbable. ¿Y a qué responde este principio? Simplemente, a la idea de que “menos es más”, a la posibilidad de expresar más conceptos con el menor número de palabras posible. Buscar una intencionalidad de carácter machista en el uso de la susodicha norma responde a concepciones tan pueriles como capciosas. Algo que la propia Academia explica muy bien en este enlace.
Puede que los partidarios de este despropósito lingüístico no hayan reparado en que, más allá de dotarla de ese supuesto carácter inclusivo, su propuesta resulta tan excluyente como divisoria, pues, decir “los trabajadores y las trabajadoras” en vez de “los trabajadores” – todos, ellas y ellos -, hace una distinción innecesaria en cuanto al género morfológico y en lo que a los sexos se refiere. Que las lenguas son algo vivo, que evolucionan con el devenir del tiempo en un proceso en el que se acuña tantas palabras nuevas como otras caen en desuso, es innegable. Pero, al mismo tiempo, las normas gramaticales y las reales academias están ahí para garantizar la pervivencia de las mismas. Que una serie de personajillos, con independencia de su color político o de su trasfondo sociocultural, venga a decirnos y a imponernos su visión particular de cómo hemos de usar nuestra lengua, entre otras cuestiones, responde más al “1984” de Orwell que a la concepción de un Estado que se autodenomina “democrático”.
En este espacio, no nos vamos a prestar a ello, ténganlo ustedes por seguro.
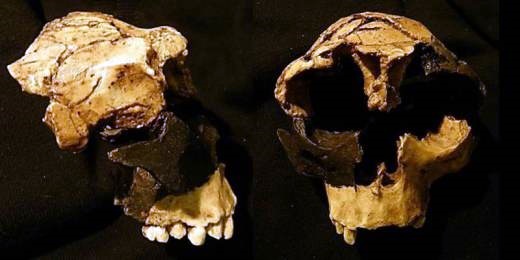
Homo antecessor. Composición de ATD6-15 y ATD6-69. Fotografía de Roberto Sáez
Hace casi veinticinco años que el equipo de Atapuerca, liderado por los paleoantropólogos Juan Luis Arsuaga, Jose María Bermúdez de Castro y Eudald Carbonell, anunciaba oficialmente el descubrimiento de una nueva especie en el genero Homo a la que denominaron ‘antecessor’, ‘pionero’, término que Bermúdez de Castro escogió al rememorar las avanzadillas que el Imperio Romano enviaba a tierras ignotas. La noticia revolucionó a la comunidad científica y puso al yacimiento burgalés en la primera línea de la investigación paleoantropológica mundial. Sin embargo, la juventud del espécimen ATD6-69, base de sus investigaciones, pronto llevaría a otros colegas de profesión a plantearse serias dudas acerca de la viabilidad de antecessor como nueva especie. Y surgió la pregunta, ¿existió realmente Homo antecessor?



















 Gaspar de Portolá (1717 – 1784)
Gaspar de Portolá (1717 – 1784)