Hace casi veinticinco años que el equipo de Atapuerca, liderado por los paleoantropólogos Juan Luis Arsuaga, Jose María Bermúdez de Castro y Eudald Carbonell, anunciaba oficialmente el descubrimiento de una nueva especie en el genero Homo a la que denominaron ‘antecessor’, ‘pionero’, término que Bermúdez de Castro escogió al rememorar las avanzadillas que el Imperio Romano enviaba a tierras ignotas. La noticia revolucionó a la comunidad científica y puso al yacimiento burgalés en la primera línea de la investigación paleoantropológica mundial. Sin embargo, la juventud del espécimen ATD6-69, base de sus investigaciones, pronto llevaría a otros colegas de profesión a plantearse serias dudas acerca de la viabilidad de antecessor como nueva especie. Y surgió la pregunta, ¿existió realmente Homo antecessor?
“Las afinidades de Homo antecessor – Una revisión de las características craneofaciales y su validez taxonómica” es un gran trabajo de investigación que da respuesta a esa incógnita. Antes de nada, queremos agradecer a nuestro amigo el Doctor en Paleoantropología D. Francesc Ribot Trafí que nos haya permitido traducirlo y publicarlo en el blog. Su ayuda y sus consejos han sido vitales para conseguir la precisión que este tipo de artículos requiere, dadas las dificultades que plantea a los que somos profanos en esa materia. Como es costumbre, hemos incorporado un glosario y unas imágenes, a fin de clarificar en la medida de lo posible todos los conceptos que definen las características anatómicas descritas en el trabajo. Y, por supuesto, el enlace a la publicación original en inglés, versión que incluye una quinta tabla que aquí no hemos reproducido por cuestiones de formato.
Sin más dilación, vamos ya con la respuesta a la pregunta que hemos planteado: ¿Existió Homo antecessor?
Las afinidades de Homo antecessor – Una revisión de las características craneofaciales y su validez taxonómica
- Francesc Ribot Trafí (Museo Municipal de Prehistoria y Paleontología Dr. Gibert, Granada, España)
- Mario García Bartual (Museo Paleontológico de Elche y Fundación Cidaris, Alicante, España)
- Qian Wang (Departament of Biomedical Sciences, Texas A&M, University College of Dentistry, Dallas, U.S.A.)
Resumen: Las afinidades filogenéticas de Homo antecessor, un hominino del Pleistoceno medio europeo, aún están por dilucidar. En este estudio, hemos procedido a una revisión exhaustiva del hipodigma TD6 en el contexto del desarrollo histórico de cuestiones paleoantropológicas que conciernen a esta especie. De acuerdo con todas las características craneofaciales de las que disponemos hasta la fecha, Homo antecessor muestra una morfología del tercio facial medio muy similar a la de especímenes catalogados como Homo erectus clásico, lo que sugiere que Homo antecessor es la variante geográfica europea de Homo erectus clásico.
Palabras clave: Atapuerca, Primeros Homo, Homo erectus, Homo antecessor, Morfología del tercio facial medio, Pleistoceno medio.
INTRODUCCIÓN
En 1997, Bermúdez de Castro y su equipo definieron una nueva especie, Homo antecessor (Bermúdez de Castro et al., 1997), basándose en los restos dentales y craneofaciales del espécimen ATD6-69, un individuo juvenil encontrado en el nivel TD6 de Gran Dolina (Atapuerca, Burgos, España), que fue datado entre 0,78 Ma (Carbonell et al., 1995) y 0,9 Ma (Parés et al., 2013).
Las características de la morfología facial de ATD6-69 permitieron a Bermúdez de Castro y sus colegas definir esta nueva especie y situarla, también, como un ancestro común de Homo neanderthalensis y Homo sapiens (ver “Diagnóstico diferencial de Homo antecessor”). Sin embargo, el estatus filogenético de H. antecessor no ha sido aceptado por unanimidad, ya que algunos autores albergan dudas de su validez como un taxón único (p. ej., Delson, 1997; Wang, 1998; Wang y Tobias, 2000a; Rightmire, 2001, 2007; Stringer, 2002; Etler, 2004; Rabadà, 2005; Ribot et al., 2006, 2007, 2017, 2018; Cartmill y Smith, 2009; Ribot y García Bartual, 2016).
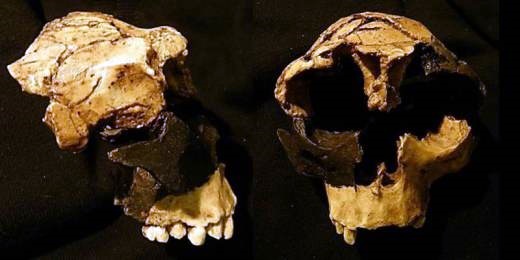
Uno de los problemas principales a la hora de aceptar a esta nueva especie humana es su diagnóstico, realizado sobre la base de un individuo no adulto. Rightmire (1998a) apuntó que la atribución a una nueva especie basada únicamente en la morfología facial de un espécimen en desarrollo resulta problemática, al tiempo que se cuestionó si la presencia de una verdadera fosa canina en un individuo joven —una de las características diagnosticadas de este taxón— es una característica morfológica apropiada para el diagnóstico de una nueva especie. De hecho, el fragmento zigomaticomaxilar ATD6-58 de Atapuerca —atribuido a un individuo adulto— muestra una fosa canina reducida que difiere de aquella del fragmento ATD6-69, atribuido a un individuo no adulto (Bermúdez de Castro et al., 1997; Arsuaga et al., 1999) (fig. 1).
En 2003, la mandíbula de un ejemplar adulto de H. antecessor fue recuperada (ATD6-96) (Carbonell et al., 2005). Del estudio de este fósil los autores concluyeron que “[…] ninguna de las características consideradas apomórficas durante el Pleistoceno medio y superior europeo del linaje Homo están presentes en ATD6-96. Una evidencia que refuerza la identidad taxonómica de H. antecessor y que resulta consistente con la hipótesis de una estrecha relación entre esta especie y Homo sapiens” (Carbonell et al. 2005. p. 5674).
Pero no queda claro que H. antecessor represente un nuevo taxón basado en características diagnósticas, en el estudio de Carbonell et al. (2005) comparan catorce características mandibulares de la mandíbula ATD6-96 con las propias de diferentes taxones de Homo (tabla 4, en Carbonell et al., 2005). La interpretación de la tabla muestra que la mayor parte de las presentes en ATD6-96 se encuentran, también, en Homo erectus clásico (ver: Anotaciones taxonómicas). De hecho, solo uno de los rasgos puede ser considerado como derivado, caso de la fosa subalveolar posterior, ahuecada. Una característica poco acusada en ATD6-96 y en algunos Homo heidelbergensis; superficial en la mayor parte de individuos de Homo erectus clásico; profunda en Homo erectus clásico de Zhoukoudian; imperceptible en Homo neanderthalensis y ciertos H. heidelbergensis. Sin embargo, la distinción esgrimida por Carbonell et al. (2005) entre una fosa subalveolar superficial y una fosa subalveolar ligeramente ahuecada es muy subjetiva, y los fósiles asignados a cualquiera de las dos categorías son virtualmente idénticos. Es más, en una revisión reciente de H. antecessor, Bermúdez de Castro et al. (2017) no otorgan rasgo derivado alguno en la mandíbula de aquél en comparación con las de humanos modernos (ver tabla 2 en Bermúdez de Castro et al. 2017).

Figura 1. Comparación entre ATD6-69, joven (derecha), y ATD6-58, adulto (izquierda). Puede observarse cómo la fosa canina en ATD6-58 es más superficial en ATD6-69. Fotografía de ATD6-69 cortesía de Roberto Sáez; fotografía de ATD6-58 tomada de Schwartz y Tattersall (2002)
También ha sido motivo de discusión que la supuesta relación ancestro-descendiente entre H. antecessor y H. sapiens y el reconocimiento de aquél como un nuevo taxón vengan reforzados por el patrón de remodelado del hueso facial de ATD6-69, que muestra una enorme semejanza con el de H. sapiens (Lacruz et al., 2013). En ambas especies, se observa un retraimiento de la región subnasal (rostro ortognático) asociado a la presencia de zonas de reabsorción alrededor del clivus nasomaxilar (Lacruz et al., 2013, 2015). Este patrón contrasta con el de KNM-WT 15000 —individuo subadulto de un H. erectus primitivo (u H. ergaster) oriundo de Turkana Occidental, en Kenia (ver las Anotaciones taxonómicas), con una datación aproximada de 1,7 Ma—, que muestra el patrón de remodelado de hueso facial primitivo observado en australopitécidos y primeros Homo, con presencia de zonas de deposición en la región subnasal, lo que provoca prognatismo facial (Bromage, 1989; Lacruz et al., 2013, 2015). De este modo, Lacruz y sus colaboradores (2013) llegaron a la conclusión de que la retracción del plano facial medio en ATD6-69, como ocurre en H. sapiens, es un aspecto clave de H. antecessor en su reconocimiento como un nuevo taxón. Sin embargo, la validez de esta característica a la hora de establecer relaciones taxonómicas resulta problemática si consideramos los hallazgos de estudios previos (Kurihara et al., 1980; McCollum, 2008).
En relación con el origen de H. antecessor, los científicos que estudian los materiales de Atapuerca han propuesto dos posibilidades diferentes: la primera sostiene que este taxón tuvo un origen africano en el que H. antecessor y H. erectus clásico comparten un ancestro común con los H. erectus primitivos (u H. ergaster) (Bermúdez de Castro et al., 1997, 2017; Arsuaga et al., 1999; Rosas, 2000).
La segunda afirma que H. antecessor tiene un origen asiático, por el cual este taxón sería el descendiente de H. erectus (Carbonell et al., 2005; Martinón-Torres et al., 2007), basada en las tremendas similitudes entre H. antecessor y H. erectus clásico de Asia en sus mandíbulas y en su morfología del tercio facial medio, así como la presencia en ambos de un tubérculo zigomaticomaxilar (Carbonell et al., 2005). Además, estos homininos presentan similitudes en cuanto a ciertas características dentales tales como la “robustez morfológica” de la dentición anterior (en forma de pala triangular, por ejemplo) y de la posterior, mediante la simplificación de la morfología oclusal, la enorme frecuencia de patrones de surco no-Y, la presencia de la cresta del trigónido medio, la fóvea anterior y la cresta transversal en los premolares (Martinón-Torres et al., 2007).
DIAGNÓSTICO DIFERENCIAL DE HOMO ANTECESSOR
El surgimiento de la nueva especie, H. antecessor, y su relación con H. sapiens se ha basado en una serie de características de ATD6-69 (Bermúdez de Castro et al., 1997) que fueron consideradas como rasgos derivados con respecto a los Homo primitivos (Homo rudolfensis, Homo habilis, H. erectus primitivos — u H. ergaster— y H. erectus clásico) y a la aparición posterior de diferentes Homo europeos (H. heidelbergensis y H. neanderthalensis). Se puso especial énfasis en la morfología del tercio facial medio de ATD6-69 y su complejo funcional nasomaxilar asociado (Bermúdez de Castro et al., 1997, 2017; Arsuaga et al., 1999), incluyendo la orientación coronal de la superficie infraorbital con la inclinación inferoposterior de su plano; la presencia de fosa canina; la inflexión maxilar horizontalmente orientada y una proyección de los huesos de la nariz; cresta zigomaticoalveolar con una raíz zigomática en una posición elevada, más la presencia de incisura malaris; crestas nasales laterales y espinales que casi convergen para formar un borde nasal inferior; la presencia de un tubérculo zigomaxilar; un moderno patrón de remodelado de los huesos de la cara y un borde superior convexo de la escama temporal. Esta combinación morfológica del tercio facial medio y craneal encontrada en los homininos de ATD6 es considerada una moderna sinapomorfia humana que formó parte por primera vez del registro fósil con la aparición de H. antecessor (tabla 1, fig. 2).
Tabla 1. Características mandibulares y craneofaciales supuestamente derivadas que definen a Homo antecessor* y son objeto de discusión en el presente estudio (ver también fig. 1)
Características diagnósticas de ‘Homo antecessor’ objeto de discusión en este estudio:
- Orientación coronal de la superficie infraorbital con inclinación inferoposterior de su plano
- Presencia de fosa canina
- Inflexión maxilar horizontalmente orientada y proyección de los huesos de la nariz
- Cresta zigomaticoalveolar arqueada con incisura malaris
- Crestas nasales laterales y espinales que casi se fusionan para formar un borde nasal inferior
- Presencia de un tubérculo zigomaxilar
- Moderno patrón de remodelado de los huesos de la cara
- Borde superior convexo de la escama temporal
*Bermúdez de Castro et al., 1997; Arsuaga et al., 1999; Carbonell et al., 2005; Bermúdez de Castro et al., 2017.

Figura 2. ATD6-69 características faciales consideradas exclusivamente derivadas con respecto a Homo sapiens y empleadas para definir la especie Homo antecessor (Bermúdez de Castro et al., 1997, 2017; Arsuaga et al., 1999): a) orientación coronal de la superficie infraorbital con presencia de una auténtica fosa canina; b) cresta zigomaticoalveolar horizontalmente arqueada, con una pronunciada raíz zigomática y un tubérculo zigomaxilar; c) inflexión maxilar determinada por la lámina infraorbital y el tabique nasal lateral (área marcada en líneas verdes); d) crestas nasales laterales y espinales muy próximas entre sí y que casi se fusionan formando el borde nasal inferior, y un moderno de patrón de remodelado de los huesos de la cara: (+) zonas de deposición; (–) zonas de reabsorción. Modificado de Schwartz y Tattersall (2002) y Lacruz et al., (2013)
MATERIAL Y MÉTODOS
Material
Entre los materiales empleados para este estudio se incluye moldes, imágenes y descripciones de los primeros Homo y H. erectus clásico de Asia (penecontemporáneo) que muestran restos del tercio facial medio, parciales o completos. Hemos estudiado, especialmente, los siguientes moldes: KNM-ER 1470; KNM-ER 1813; OH 24; OH 13; KNM-ER 3883; KNM-ER 3733; KNM-WT 15000; Gongwangling; Sangiran 17; Zhoukoudian (reconstrucción de Weidenreich —1937a— y Sawyer y Tattersall —1995 [y Tattersall y Sawyer, 1996]—). También, una muestra de 35 cráneos humanos del Laboratorio de Paleopatología y Paleoantropología del Museo Nacional de Arqueología de Cataluña en Barcelona. En cuanto al estudio de los restos de H. antecessor, se utilizó una réplica de gran calidad de ATD6-69 (no el molde estándar presente en la mayoría de los laboratorios), traído por los investigadores del grupo de Atapuerca para una exposición especial en el Instituto de Paleontología Miquel Crusafont de Sabadell en Barcelona. Uno de nosotros (F.R.T.) tuvo acceso a esta pieza durante meses, y se le permitió estudiarla y compararla con las imágenes y descripciones de ATD6-69 que el equipo de Atapuerca había realizado. De igual manera, hemos utilizado las fotografías y las descripciones originales del material.
Anotaciones taxonómicas
En este estudio, la morfología del tercio facial medio de H. antecessor se compara con la propia de las especies de Homo más antiguas (primeros Homo y H. erectus primitivos [o H. ergaster]) y con las especies penecontemporáneas de H. erectus clásico. Los taxones empleados y los especímenes que los representan son los siguientes:
1. Primeros Homo
Hemos considerado como tales a aquellos especímenes datados entre ~2,4 y 1,4 Ma (tabla 2). Aunque la consideración de primeros Homo es controvertida, en este trabajo hemos optado por una clasificación más clásica y, de acuerdo con Antón et al. (2014), distinguido entre los Grupos 1470 y 1813, denominados H. rudolfensis y H. habilis, respectivamente, por varios autores como, por ejemplo, Wood, 1992; Kimbel et al., 1997; Strait et al., 1997; Prat, 2004; Kimbel, 2009; Schrenk et al., 2015; Simpson, 2015.
Tabla 2. Especímenes fósiles clasificados aquí como primeros Homo
Espécimen Edad(Ma) Autor
A.L. 666-1 2,33 Kimbel et al., 1997
Grupo 1470
KNM-ER 1470 2,03-2,09 Joordens et al., 2013
KNM-ER 62000 1,95-1,98 Joordens et al., 2013
Grupo 1813
OH 24 1,88 Hay, 1976
KNM-ER 1813 ~1,86 Feibel et al., 2009
OH 62 1,75-1,85 Johanson et al., 1987
OH 65 ~1,78 Blumenschine et al., 2003
OH 13 1,60 Hay, 1976
KNM-ER 42703 1,44 Spoor et al., 2007
En el Grupo 1470 incluimos los especímenes KNM-ER 1470 y KNM-ER 62000. Leakey et al. (2012) incluyen KNM-ER 1470 y KNM-ER 62000 en el mismo grupo a causa de sus similitudes morfológicas. En el Grupo 1813, incluimos los especímenes OH 24, OH 62, OH 65, KNM-ER 1813 y KNM-ER 42703.
La clasificación del maxilar OH 65 resulta controvertida: Blumenschine et al. (2003) y Clarke (2012) la agrupan con KNM-ER 1470, y Clarke (2012) separa KNM-ER 1470 y OH 65 (H. habilis) de KNM-ER 1813 y OH 24 (Australopithecus cf. africanus). Sin embargo, la inclusión de KNM-ER 1470 y OH 65 en el mismo grupo ha sido cuestionada: Spoor et al. (2007) señalan que la morfología del maxilar KNM-ER 42703 está muy próxima a la de los grandes especímenes de H. habilis (OH 65 y KNM-ER 1805), y es muy diferente de la de KNM-ER 1470. Además, Leakey et al. (2012) y Antón (2012) distinguen entre OH 65 y KNM-ER 1470 y KNM-ER 62000, ya que OH 65 se muestra mucho más prognática a nivel subnasal y presenta las raíces zigomáticas más posteriores, el clivus nasoalveolar (subnasal) está curvado en el margen alveolar y sus alveolos no forman parte de la dentadura anterior; los incisivos se proyectan más allá de la línea canina y el paladar se dispone parabólicamente (como ocurre con KNM-ER 1813 y a diferencia de la forma en U de KNM-ER 62000). Es por ello que incluimos a OH 65 en el Grupo 1813.
Otro espécimen de difícil clasificación es KNM-ER 1805. Muchos autores lo adscriben a los primeros Homo (entre ellos, Wood 1991, 1992; Kimbel et al. 1984, 1997, 2004; Wood and Richmond 2000, Williams et al. 2012; Schrenk et al. 2015); sin embargo, están quienes lo incluyen entre los australopitecos: Falk (1986) señala que el patrón del surco cortical de ER 1805 presenta afinidades con Australopithecus; Tobias (1980) adscribió el espécimen a Paranthropus boisei sobre la base de su morfología molar; de igual manera, Prat (2002, 2004), basándose en un análisis cladístico, en el que consideró 122 características craneales, concluye que ER 1805 debería ser incluido en el género Paranthropus. Finalmente, Grossman (2009) señala que ER 1805 ni presenta la morfología típica de Homo ni la de Paranthropus, llegando a la conclusión que la serie de rasgos que KNM-ER 1805 presenta es ciertamente única. Dadas las diferentes interpretaciones sobre este fósil, hemos optado por excluir a KNM-ER 1805 de este estudio.
2. Homo erectus primitivos (u Homo ergaster)
Consideramos como los H. erectus primitivos (u H. ergaster) a aquellos homininos datados en una escala temporal aproximada de entre 1,8 a 1,3 Ma (tabla 3) —clasificados como Earliest H. erectus por Antón (2003). De los especímenes que hemos estudiado, Antón incluye en el grupo Earliest H. erectus a los especímenes africanos KNM-ER 3733 y KNM-WT 15000, restos de Dmanisi y Sangiran 4 (hemos añadido los fósiles asiáticos Gongwangling y Sangiran 17, clasificados por Antón como Early H. erectus porque presentan cronologías más recientes que Earliest H. erectus más antiguos; sin embargo, las últimas dataciones estiman los restos de Gongwangling en 1,63 Ma —Zhu et al., 2015— y los de Sangiran 17 entre 1,30 y 1,25 Ma —Larick et al., 2001, 2004; Antón et al., 2007). Pese a lo cual, Antón considera del mismo grupo a los especímenes de África Oriental, los de Dmanisi y Sangiran 4. Así mismo, dada su similar morfología, Kaifu et al. (2010) y Lordkipanidze et al. (2013), atribuyeron fósiles de África Oriental y las muestras de Dmanisi y Sangiran al mismo grupo.
Tabla 3. Especímenes fósiles aquí clasificados como Homo erectus primitivos (u Homo ergaster)
Espécimen Edad (Ma) Autor
Africa
KNM-ER 3733 1,78 Wood, 1991
KNM-ER 3883 1,50-1,65 Wood, 1991
KNM-WT 15000 1,50 Walker y Leakey, 1993
Oriente Medio (Dmanisi)
D2282 ~1,77 Gabunia et al., 2000; Rightmire et al., 2006
D2700 ~1,77 Gabunia et al., 2000; Rightmire et al., 2006
D3444 ~1,77 Gabunia et al., 2000; Rightmire et al., 2006
D4500 ~1,77 Gabunia et al., 2000; Rightmire et al., 2006
Asia
Gongwangling 1,63 Zhu et al., 2015
Sangiran 4 >1,5 Swisher et al., 1994; Antón 2003; Antón y …………………………………………………..Swisher, 2004; Larick et al. 2001, 2004; Antón et …………………………………………………. al. 2007
Sangiran 17 1,25-1,30 Larick et al. 2001, 2004; Antón et al. 2007
La dificultad de clasificar taxonómicamente estos homininos se aprecia claramente en la disparidad de interpretaciones entre autores diferentes. Así, Wang y Tobias (2000a) han empleado el término H. ergaster para homininos que presentan un patrón facial tipo I, y Homo erectus tardío (H. erectus clásico en este estudio) para homininos con un patrón facial tipo II, distinguiendo con claridad los modelos faciales entre ambos grupos.
Por otra parte, Williams et al. (2012) consideran como H. erectus (u H. ergaster) los restos de África Oriental. Recientemente, Tattersall (2015) ha venido a demostrar que es necesario separar a H. ergaster de H. erectus, y subraya la gran variedad de clasificaciones para los restos de Dmanisi: H. erectus (Gabunia y Vekua, 1995; Henke, 1995; Bräuer y Schultz, 1996; Vekua et al., 2002); H. ergaster (Gabunia et al., 2000); H. georgicus (Gabunia et al., 2002); H. erectus georgicus (Rightmire et al., 2006), y H. erectus ergaster georgicus (Lordkipanidze et al., 2013). Hemos optado por una denominación que combine la empleada por Antón (2003) y la manejada por Williams et al (2012), de forma que clasificamos estos restos como H. erectus primitivos (u H. ergaster).
Este grupo presenta una morfología facial que lo distingue tanto de los primeros Homo y de H. erectus clásico como de H. sapiens. Morfología que se caracteriza por:
- ausencia de fosa canina y presencia de surco maxilar
- cresta zigomaticoalveolar oblicua o recta
- ausencia de incisura maxilaris
- huesos de la nariz poco proyectados
- inflexión maxilar débil
- raíz zigomática adelantada con respecto al margen lateral nasal
- origen bajo del hueso zigomático (o malar)
- borde superior rectilíneo de la escama temporal
3. Homo erectus clásico
Los especímenes de H. erectus clásico incluidos en este estudio están datados entre ~1 Ma y ~0,6 Ma (homininos clasificados como Middle H. erectus por Antón, 2003 [Zhoukoudian y Nanjing], a los que hemos añadido los cráneos de Yuxian). En este grupo, hemos incluido, también, los especímenes africanos KNM-OL 45500, OH 12 y BOU-VP-2/66, aunque no sean parte de esta investigación, ya que KNM-OL 45500 es un fragmento frontal y temporal, OH 12 está muy troceado y BOU-VP-2/66 es una calota craneal (tabla 4).
Características anatómicas del tercio facial medio
El término mediofacial se aplica a la porción del cráneo anterior que es visible in norma frontalis, vista de frente, y que está verticalmente delimitada por la distancia que va desde el plano alveolar de la mandíbula hasta la intersección entre el extremo superior de la nariz con el arco frontal (nasion), y en sentido transversal de cigión a cigión (Pope 1991). La topografía de esta zona difiere en los diferentes grupos de homininos, pero, en H. sapiens se presenta enormemente curvada. El rostro flexionado de H. sapiens viene determinado por la orientación coronal de la placa ósea infraorbital respecto de la superficie inferior que se inclina hacia abajo y ligeramente hacia atrás (fosa canina), y por la dirección más sagital del tabique lateral nasal. A lo largo de la articulación de estas dos superficies maxilares, se forma una zona de flexión. De igual manera, la cresta zigomaticoalveolar aparece generalmente curvada y a menudo presenta una incisura malaris (Rak, 1983, 1986; Arsuaga et al., 1999).
Para este estudio, comparamos las características descritas como propias en H. antecessor, del tercio facial medio y en número de seis, más el patrón de remodelado del hueso facial —referido la flexión mediofacial— y la forma del borde superior de la escama temporal. Es más, los comparamos con aquellos de los homininos más ancestrales y penecontemporáneos (primeros Homo, H. erectus primitivos [u H. ergaster] y H. erectus clásico).
Orientación coronal de la superficie infraorbital.
Inflexión maxilar. Esta inflexión viene determinada por la superposición del tabique lateral nasal y la inclinación hacia abajo y ligeramente hacia atrás de la placa infraorbital (Bermúdez de Castro et al., 1997; Arsuaga et al., 1999). Arsuaga et al. (1999) añaden también a esta morfología la proyección de los huesos de la nariz, posición adelantada del límite superior del borde nasal.
Table 4. Especímenes fósiles clasificados aquí como Homo erectus clásico
Espécimen Edad (Ma) Autor
Africa
BOU-VP-2/66 ~1 Asfaw et al., 2002
KNM-OL 45500 0,9-0,97 Potts et al., 2004
OH 12 0,78 Tamrat et al., 1995; Antón, 2004
Asia
Yunxian 0.8-~0.94 Yan, 1993; Feng, 2008; Feng et al, 2011;
…………………………………………… De Lumley y Li, 2008
Zhoukoudian* ~0,8 Shen y Jin, 1991; Shen 2001;
……………………………………………Shen et al. 2009
Tangshan (Nanjing) ~0,62 Zhao et al., 2001
* Estrato inferior de la Localidad 1
Fosa canina. La fosa canina se define como una depresión infraorbital que incluye la mayor parte del proceso zigomático de la mandíbula (Mellinger, 1940). Algunos autores añaden a esta definición el hecho de que esta depresión infraorbital produce una incurvación horizontal y una incurvación de la cresta zigomáticoalveolar (Arsuaga et al., 1999; Maddux y Franciscus, 2009). Esta descripción coincide con lo que Maureille y Houët (1997) llaman depresión infraorbital. Sin embargo, Arsuaga et al. (1996, 1999) sugieren lo contrario cuando describen el fragmento maxilar AT-404 de Sima de los Huesos (Atapuerca) (fig. 4 en Arsuaga et al., 1996, y fig. 7 en Arsuaga et al., 1999), un posible H. heidelbergensis, con presencia de fosa canina y cresta zigomaticoalveolar recta y oblicua. Así mismo, la misma morfología de AT-404 se observa en el cráneo del probable H. heidelbergensis de Steinheim y en algunos especímenes modernos (fig. 3).

Figura 3. Ejemplos de fosa canina con cresta zigomaticoalveolar recta-oblicua: A) Australopithecus sediba; B) probable H. heidelbergensis, Steinheim (imagen revertida); C) probable H. heidelbergensis, AT-404, Atapuerca (obtenida de Arsuaga et al., 1999); D) Humano moderno (obtenida de Stan et al., 2013)
Cresta zigomaticoalveolar. Esta cresta forma el límite inferior del proceso zigomático del maxilar y se eleva desde la pared lateral del maxilar. La cresta zigomaticoalveolar puede ser observada mejor desde una vista frontal, y determinada por la línea que se desplaza desde el plano alveolar al zigomaxilar (Pope, 1991; Koesbardiati, 2000). En este estudio, aplicamos las categorías definidas por Etler (1994): oblicuo (recto y oblicuamente orientada), arqueada y horizontal. Desde una vista frontal, la cresta zigomaticoalveolar está sujeta al desarrollo de la incisura malaris y de las estructuras relacionadas con el tubérculo malar (Rak, 1983). La incisura malaris puede estar presente o no (fig. 4)

Figura 4. Caras de humanos modernos con cresta zigomaticoalveolar curvada A) ausencia de incisura malar; B) con incisura malar. Colección del Laboratorio de Paleopatología y Paleoantropología, Museo Nacional de Arqueología de Cataluña en Barcelona. Fotografías tomadas por F.R.T. con la autorización del Dr. Domènec Campillo.
Margen nasal inferior. Tres crestas están implicadas en la formación del margen nasal inferior: 1) espinal —se origina en la espina nasal anterior—; una cresta espinal puede aparecer localizada en el plano posterior, a lo largo del polo posterior del proceso alveolar, o en el plano anterior, a lo largo del clivus nasoalveolar (o clivus subnasal); 2) lateral —se origina en el margen lateral de la abertura piriforme, y 3) turbinal —surge junto al extremo anterior del cornete nasal (Gower, 1923). El margen inferior nasal puede formarse por la presencia de una única cresta o por la combinación de dos o tres de ellas, dando lugar a hasta seis categorías (Gower, 1923; Lahr, 1994; Franciscus, 2003) (la ausencia de margen nasal inferior es considerada como la séptima categoría). La espina nasal, mencionada anteriormente, es la extensión anterior de la cresta de los incisivos que da lugar a un proceso espinal que se proyecta hacia el proceso alveolar. Los hominoideos y muchos homininos fósiles no presentan esta extensión anterior, por lo tanto, no poseen una auténtica espina nasal anterior, aunque generalmente exhiben una pequeña protuberancia o tubérculo en el clivus nasoalveolar en la unión más anterior del tabique nasal (septum).
Tubérculo zigomaxilar. Este tubérculo ocupa la sección inferior de la cara anterior del hueso zigomático, que marca el nacimiento anterior del músculo mandibular (Rightmire 1998b). Hemos de distinguirlo de la tuberosidad malar, que es una elevación relativamente prominente sobre la superficie malar paralela al extremo inferior del hueso, situada entre los márgenes orbital y exento del maxilar (Koesbardiati 2000).
Patrón de remodelado de los huesos de la cara. El remodelado óseo es uno de los factores fundamentales del proceso del crecimiento craneofacial (McCollum, 2008). El remodelado óseo facial se refiere a diferentes combinaciones de deposición y reabsorción de hueso vivo en las superficies interior y exterior de una lámina ósea (McCollum, 2008). La presencia de áreas de reabsorción de hueso en el maxilar y en el clivus nasoalveolar durante la ontogenia se asocia con un rostro ortognático, mientras que la aparición de zonas de deposición en el hueso en el maxilar y en el clivus nasoalveolar contribuye a la formación de un rostro prognático (Enlow and Wang, 1965; Enlow, 1966; Bromage, 1989; McCollum, 2008)
Borde superior de la escama temporal. El borde superior de la escama temporal puede presentar dos morfologías: plana o rectilínea, asociada a una capacidad craneal limitada; y convexa o arqueada, asociada a una capacidad craneal aumentada.
RESULTADOS
Primeros Homo
La morfología facial de los primeros Homo presenta dos morfotipos posibles: el que aparece en el Grupo 1470, en primer lugar, y el presente en el Grupo 1813, en segundo lugar. Sin embargo, previo a estos dos grupos, nos encontramos con el maxilar A.L. 666-1 de Hadar, datado en unos 2.4 Ma y clasificado como Homo aff. H. habilis (Kimbel et al., 1997). La configuración de este maxilar podría revelar el patrón morfológico presente en los primeros Homo. Kimbel et al. (1997) describen este fósil como poseedor de un clivus nasoalveolar que no resulta muy prognático, que presenta corrugaciones en el alvéolo de los incisivos centrales; con una ligera protrusión de los márgenes nasales laterales; una cresta nasal lateral que se extiende y se curva medianamente sobre el clivus; un tubérculo espinal (la espina nasal no se observa en vista lateral); una cresta espinal que marca el margen nasal inferior, y una depresión superficial que da forma a una fosa canina rudimentaria en forma de lágrima.
Grupo 1470
La primera morfología, caracterizada por KNM-ER 1470 y KNM-ER 62000, muestra un plano infraorbital anteroinferiormente orientado, plano y coronal, con la separación entre la superficie maxilar anterior y la raíz del proceso zigomático pequeña (plano coronal comprimido), sin fosa canina y con una cresta zigomaticoalveolar con un origen bajo y adelantado (sobre P3/P4 en KNM-ER 62000 y sobre P4 en KNM-ER 1470 —Leakey et al., 2012), y con una cresta zigomaticoalveolar recta y diagonalmente orientada; eversión lateronasal muy poco acentuada (escasa orientación sagital del tabique lateral nasal); plano subnasal ortognático; orificio infraorbital próximo al margen orbital; clivus nasoalveolar aplastado y de gran anchura, que se curva hacia atrás alrededor de las raíces P3 (eminencia de P3 bien definida), los alvéolos caninos son parte integral de la fila anterior; paladar de gran anchura y profundidad; parte superior del rostro hasta cierto punto estrecha, y un seno maxilar que se desarrolla hacia adelante hasta el plano coronal del margen nasal inferior. En KNM-ER 1470, el margen nasal inferior está formado por el desarrollo lateral de la cresta espinal (conformación tipo 2 del margen nasal —Franciscus 2003). Finalmente, en KNM-ER 1470, el límite superior de la escama temporal es rectilíneo.
Grupo 1813
Este grupo incluye a KNM-ER 1813, KNM-ER 42703, OH 24, OH 65 y OH62, y presenta un modelo facial diferente al del anterior: el plano infraorbital está orientado coronalmente con inclinación posteroinferior, al tiempo que presenta una mayor superficie maxilar lateral (sin compresión) que el grupo anterior, dando lugar a una fosa canina extensa y profunda (Rak, 1983; Kimbel et al., 1997; Schwartz and Tattersall, 2003); la raíz zigomática se sitúa posteriormente del plano coronal (sobre P4/M1 en todos los especímenes de este grupo —Leakey et al., 2012), con un origen más elevado; la cresta zigomaticoalveolar es arqueada (KNM-ER 1813, KNM-ER 42703, OH 24), o recta y orientada oblicuamente (OH 62, pero con una fosa canina muy superficial —Schwartz y Tattersall, 2003); además, no presenta incisura malaris ni tubérculo zigomaxilar (Pope, 1991); el desplazamiento hacia adelante del seno maxilar hasta el plano coronal del margen nasal inferior no se hace evidente en el Grupo 1813; pero sí hay presencia de inflexión maxilar y eversión lateral nasal (acentuada en KNM-ER 1813 y, en cierto modo, más atenuada en OH24); la zona subnasal es estrecha y prognática; el margen nasal inferior está muy poco desarrollado: en KNM-ER 1813 y OH 24 está formado por el desarrollo lateral de una pequeña cresta espinal redondeada que no se une con la cresta lateral (categoría 2 de Franciscus 2003), al tiempo que la cresta lateral desciende por clivus nasoalveolar, mientras que en OH 62 la cresta espinal se desplaza lateralmente hasta fusionarse con la cresta turbinal, ambas separadas de la cresta lateral (categoría 3 de Franciscus 2003). En la mayoría de los primeros Homo, solo aparece un pequeño tubérculo en la línea media del margen nasal inferior en vez de una espina nasal anterior. Las excepciones son OH 24 (Rightmire 1993; Franciscus y Trinkaus 1988; McCollum et al. 1993) y OH 62 (Johanson et al. 1987), pues, ambos muestran una espina nasal anterior bien definida. KNM-ER 1813 presenta una pequeña protrusión (elevación) a cada lado de la línea media que sugiere la existencia in vivo de una espina nasal anterior (Schwartz y Tattersall, 2003). La espina nasal en OH 24 y en OH 62 no es observable desde el aspecto lateral.
En OH 13 y SK 27, el patrón de remodelado y crecimiento del hueso facial muestra una fuerte deposición de hueso en la cara anterior similar a la encontrada en australopitécidos y relacionada, posiblemente, con prognatismo nasoalveolar (Bromage, 1989). La morfología del borde superior de la escama temporal es variable: rectilínea en OH 24 o curva (convexa) en KNM-ER 1813.
Homo erectus más antiguos (u Homo ergaster)
En H. erectus primitivos (u H. ergaster), encontramos dos morfologías del tercio facial medio relacionadas con su etapa de desarrollo vital (Wang y Tobias, 2000a). La morfología que presentan los individuos inmaduros (KNM-WT 15000 y D2700) se caracteriza por: orientación de la región infraorbital en el plano coronal, pero no está inclinada hacia adelante ni hacia atrás (uniplanar); huesos nasales proyectados y maxilar orientado sagitalmente en paralelo a la abertura piriforme, y una débil inflexión maxilar. El proceso zigomático de la mandíbula es plano, y presenta un surco maxilar (desaparición parcial de la fosa canina, que se transforma en un surco que desciende desde el conducto (foramen) infraorbital. La cresta zigomaticoalveolar está arqueada en D2700 y oblicua en KNM-WT 15000, sin incisura malaris y con la ausencia de un tubérculo maxilar.
El margen nasal inferior en KNM-WT 15000 (Rightmire, 1998b) está formado por la cresta espinal, que se desplaza lateralmente hacia la cresta lateral, pero sin fusionarse, ya que la cresta espinal se encuentra a escasos milímetros tras el margen nasal lateral (categoría 3 de Franciscus 2003), mientras que D2700 no presenta cresta espinal. Finalmente, hay presencia de espina nasal, aunque no es visible desde una vista lateral.
La morfología facial de los especímenes adultos se caracteriza por una acusada flexión maxilar (los maxilares, en paralelo a la abertura piriforme, se desplazan hacia adelante), así como por las siguientes: un surco maxilar más que evidente; la cresta zigomaticoalveolar es arqueada u horizontalmente recta; los huesos de la nariz se proyectan hacia adelante (prominencia nasal); una cresta espinal, con un mayor o menor grado de desarrollo, da forma al margen nasal inferior (excepto en Sangiran 17). KNM-ER 3733 y Sangiran 17 exhiben un tubérculo zigomaxilar (Pope, 1991; Leakey y Walker, 1985; Rightmire, 1998b; Schwartz y Tattersall, 2003). La existencia de una espina nasal prominente y observable desde una vista lateral es visible en Sangiran 4 (Sawyer y Tattersall, 1995; Tattersall y Sawyer, 1996; Franciscus y Trinkaus, 1988; Rightmire, 1998b; McCollum, 2000; Schwartz y Tattersall, 2003) y en Gongwangling (Ju-kang, 1966; Rightmire, 1998b); mientras que el origen del proceso zigomático en todos los H. erectus primitivos (u H. ergaster) es lateral al margen lateral de la abertura nasal. Se ha observado, también, la presencia de marcadas corrugaciones en D2282 (Rightmire et al. 2006), D4500 (Rightmire et al. 2017) y Gongwangling (Rightmire 1998b).
El patrón de remodelado de crecimiento del hueso facial solo ha sido estudiado en el individuo adolescente KNM-WT 15000, que muestra un patrón primitivo, por ejemplo, un clivus liso sin arrugas o sin reabsorción alrededor de las raíces de los dientes anteriores (Lacruz et al., 2013, 2015). En todos los especímenes, el borde superior de la escama temporal es rectilíneo, excepto en D3444.
Homo erectus clásico
La morfología del tercio facial medio de este grupo, especialmente en H. erectus clásico de China (Yuxian [Li y Etler, 1992], Zhoukoudian [Wang y Tobias, 2000a; Etler, 2004], y Tangshan —Nanjing [Wang y Tobias, 2000 a,b, 2001; Liu et al., 2005]) se caracteriza por:
- placa infraorbital coronalmente orientada y posteroinferiormente inclinada
- el surco maxilar se ha transformado en fosa canina
- flexión maxilar muy pronunciada
- la cresta zigomaticoalveolar es muy arqueada, con una incisura malaris desarrollada
- situación posterior del nacimiento del proceso zigomático respecto del margen lateral nasal
- presencia de tubérculo zigomaxilar
- huesos de la nariz evertidos (nariz proyectada)
En los cráneos de Yuxian, la morfología del tercio facial medio se caracteriza por presentar:
- una placa infraorbital orientada coronalmente
- un nacimiento elevado de la raíz zigomática
- la presencia de una fosa canina
- una cresta zigomaticoalveolar recta con una marcada incisura malaris (Li y Etler, 1992)
En cuanto a la morfología del tercio facial medio, acerca del espécimen de Zhoukoudian, Wang y Tobias (2000a, p. 25) han afirmado: “Medialmente, la porción del maxilar lateral a la apertura piriforme está orientada lateralmente, mientras que la zona infraorbital lo está hacia adelante; de ese modo, conforman conjuntamente una flexión cóncava”. Y es que, efectivamente, en la nueva reconstrucción del Homo erectus clásico de Zhoukoudian (Sawyer y Tattersall, 1995; Tattersall y Sawyer, 1996; Wang, 1998), la incisura malaris se presenta menos constreñida que en la de Weidenreich (1937a), lo que resulta en una cresta zigomaticoalveolar más amplia. Este ensanchamiento da lugar a una cavidad infraorbital mucho más pronunciada. Desafortunadamente, resulta imposible determinar si esta característica es una verdadera fosa canina o un surco maxilar (como en la reconstrucción de Weidenreich), porque el maxilar empleado en esta reconstrucción está erosionado. Otros maxilares de la cueva de Zhoukoudian aparecen fragmentados, lo que dificulta la observación de la región infraorbital de los mismos (comunicación personal de Tattersall en 2016). Según Wang (1998) y Wang y Sun (2000), la reconstrucción de Weidenreich del esqueleto facial femenino de H. erectus clásico implica fragmentos craneales de individuos diferentes (de ambos sexos), una calota craneal femenina (cráneo número XI o LII) y un maxilar masculino (No V, mencionado como No II); de manera que el hueso zigomático tuvo que desplazarse lo suficiente como para unirse a los huesos maxilar y frontal. El zigoma se orienta lateralmente para encontrarse con los huesos frontal y maxilar, resultando en una orbita más grande y más baja. Sin embargo, tal y como se ha visto en el cráneo masculino reconstruido (Sawyer y Tattersall, 1995; Tattersall y Sawyer, 1996), la topografía general del esqueleto del tercio facial medio, tal y como Weidenreich (1937a) lo había reconstruido para la población de Zhoukoudian, todavía es de utilidad. En la cara de Tangshan, las superficies anterior y posterior del proceso zigomático del maxilar llegan a encontrarse en un borde redondeado. Este se extiende hacia arriba y hacia los lados, y después gira hacia abajo, formando una incisura malaris (Liu et al., 2005). Tangshan muestra, en la zona inferior de la raíz zigomática, una fosa canina muy bien desarrollada asociada a una pronunciada flexión del tercio facial medio (Wang y Tobias, 2000 a,b; 2001).
H. erectus clásico de Yunxian 2 y de la Localidad 1 de Zhoukoudian (cráneos III, V, XI, y XII) muestran una escama temporal con un borde superior convexo (Rightmire, 1998a; Etler, 2004; Wu y Poirier, 1995). A la inversa, el examen del hueso parietal de Tangshan muestra que la escama temporal estaba relativamente elevada y curvada (convexa) (Liu et al., 2005).
DISCUSIÓN
El conocimiento de los patrones de morfología y topografía faciales son importantes para comprender la variabilidad y las relaciones evolutivas de los primeros homininos (Tobias, 1967, 1991; Rak, 1983, 1986; Pope, 1991; Rightmire, 1998b, 1993; Wang y Tobias, 2000a). En el diagnóstico diferencial de H. antecessor, algunos científicos han apuntado que este taxón tiene una morfología mediofacial derivada respecto a las formas de Homo más primitivas (primeros Homo — Grupos 1470 y 1813 —, H. erectus primitivos [u H. ergaster] y H. erectus clásico), porque presenta una cara flexionada (p. ej., orientación coronal de la superficie infraorbital; nariz proyectada; inflexión maxilar — con tabique lateral de la abertura nasal evertido con respecto a la superficie infraorbital adyacente—; fosa canina, y cresta zigomaticoalveolar horizontalmente arqueada) (Bermúdez de Castro et al., 1997, 2017; Arsuaga et al., 1999; Rosas, 2000). En los hominoideos africanos actuales (Gorila y Pan), los australopitecos del este africano, el grupo 1813 y en H. sapiens, la región infrorbital se orienta coronalmente y presenta una inclinación posteroinferior que se extiende hacia la cresta zigomaticoalveolar; sobre el foramen infraorbitario y justo debajo del borde orbital inferior se encuentra el contrafuerte transversal, que forma una cresta (o rugosidad) que se extiende medialmente desde el cuerpo del hueso zigomático hacia la parte superior de la abertura nasal. Este contrafuerte transversal es más pronunciado en Gorilla, y decrece gradualmente hasta hacerse rudimentario en H. sapiens. Así, el contrafuerte transversal junto con la abertura nasal (más o menos pronunciada —muy elongado en Gorilla y discreto en H. sapiens) y la inclinación posteroinferior de la región infraorbital forman la fosa canina. En las especies cuya inclinación del plano infraorbital es anteroinferior, el contrafuerte transversal está obliterado (p. ej., Australopithecus africanus, Paranthropus, y el Grupo 1470) y la fosa canina ha sido modificada (surco maxilar en A. africanus, y fosa maxilar en P. robustus) o ha desaparecido (P. boisei y Grupo 1470). Esto podría ser debido a la compresión anteroinferior del plano coronal causada por la acusada expansión anterior del seno maxilar. Sin embargo, a este respecto, Arsuaga et al. (1999) concluyeron que la presencia de fosa canina y la cara flexionada son características derivadas en H. sapiens aparecidas por primera vez en H. antecessor y, citando a Kimbel et al. (1997), argumentaron que en todos los primeros Homo y en H. erectus primitivos (u H. ergaster) (p. ej., KNM-ER 3733 y KNM-WT 15000) se observa una compresión del plano coronal que resulta en la típica cara plana que estos homininos presentan. Pero la verdadera interpretación de Kimbel et al. al respecto es diferente, pues, estos autores señalan que el Grupo 1470 muestra una expansión notable del seno maxilar que provoca una compresión posteroanterior del plano coronal y, por lo tanto, la obliteración de la fosa canina, al tiempo que la superficie anterior del proceso zigomaticomaxilar está prácticamente en el mismo plano coronal que la abertura nasal, lo que resulta en una cara plana; mientras que en el Grupo 1813, el sinus maxilar no se extiende anteriormente que la fosa incisiva; además, desde una perspectiva lateral, el proceso zigomático maxilar y la abertura nasal se encuentran separados por un intervalo topográfico considerable, al tiempo que las raíces del proceso zigomático aparecen ubicadas posteriormente, formando de este modo la fosa canina, llegando a la conclusión de que esta última es la morfología generalizada en homininos (descripción que incluye, también, a KNM-ER 3733 ).
Por otra parte, la morfología de la cresta zigomaticoalveolar resulta igualmente compleja. Así, la curvatura general de dicha cresta y la presencia o ausencia de la incisura malaris son características interrelacionadas, cuya expresión depende de la altura del origen del masetero (Freidline et al. 2013). Especies con un bajo origen del masetero y una orientación coronal del plano infraorbital presentan una cresta zigomaticoalveolar arqueada, y pueden o no tener una incisura malaris (en el Grupo 1813 no existe, mientras que en H. erectus primitivos [u H. ergaster] este rasgo es variable, mientras que en H. erectus clásico y en H. sapiens, sí está presente). Por otra parte, en las especies que presentan un nacimiento alto del masetero y un plano infraorbital orientado coronalmente, la cresta zigomaticoalveolar es recta y se orienta oblicuamente (australopitecos y parantropos) (Freidline et al., 2013).
Las características faciales modernas de H. antecessor en realidad son retenciones ancestrales relacionadas con humanos posteriores, y pueden considerarse como parte de un patrón generalizado de arquitectura facial (Wang y Tobias, 2000a; Freidline et al., 2013; Ribot et al., 2017). Esta conclusión coincide plenamente con lo que Rak (1986) denomina un rostro generalizado, presente en muchos primates, incluyendo a humanos modernos y especies fósiles del género Homo, que se caracteriza por:
- una superficie infraorbital orientada en el plano coronal y con una inclinación posteroinferior, inclinación que forma la fosa canina en la intersección con la abertura nasal
- la cresta zigomaticoalveolar aparece curvada y se une al cuerpo del maxilar a mitad del arco dental, aproximadamente, o tras él
- clivus nasoalveolar convexo en sección transversal.
El rostro que presenta H. sapiens es, en muchos aspectos y en el sentido más básico, plesiomórfico, una característica presente, incluso — aunque muy débilmente — en los australopitecos del este africano y en los primeros Homo. Cuando cada característica del tercio facial medio es considerada de manera individualizada, o todas en conjunto, el patrón en Australopithecus afarensis responde a una morfología prototípica que podría evolucionar gracias a modificaciones relativamente simples hacia el rostro devenido en H. sapiens. La transformación completa del rostro plesiomórfico hasta el actual es un proceso gradual que culmina con la formación de un rostro más flexionado y ortognático. Así, la parte anterior de esta región del tercio facial medio se ve tremendamente afectada por las fuerzas de oclusión y de mordida de premolares y dientes anteriores, mientras que el proceso zigomático y la cresta zigomaticoalveolar se ven afectadas por la carga de los molares (Wang et al., 2010; Rodrigues Freire et al., 2014). La tensión más intensa de estas fuerzas (tensión extensiva y tensión compresiva) se produce en la fosa canina y en el proceso maxilar anterior, dos de las estructuras más relacionadas con la flexión facial. La ontogenia posnatal podría estar influenciada por factores epigenéticos, como son los biomecánicos, que pueden dilucidar diferencias funcionales entre formas faciales diferentes. En este sentido, hay que destacar que un rostro ortognático está mejor adaptado para absorber las tensiones pre y paramasticatoria que un rostro prognático (Wang et al., 2010).
Comparado con los australopitecos del este africano, los rostros del Grupo 1813 aparecen más flexionados, con tabiques más evertidos y adyacentes al margen nasal lateral y a una región infraorbital más inclinada posteroinferiormente, que resultan en fosas caninas menos restringidas (más expandidas) y menos profundas. Esta tendencia creciente de la flexión del tercio facial medio prosigue en H. erectus primitivo (u H. ergaster) (donde la fosa canina ha sido parcialmente obliterada y se ha transformado en el surco maxilar), para culminar en H. erectus clásico, en el que la combinación de una mayor inclinación posteroinferior del plano infraorbital orientado coronalmente, una cresta zigomaticoalveolar arqueada con incisura malaris y una mayor eversión del tabique adyacente al margen lateral nasal van unidos a la presencia de una “auténtica” fosa canina (fig. 5).
Esta morfología del tercio facial medio presente en H. erectus clásico es similar a la que encontramos en H. sapiens, e idéntica a la que aparece en H. antecessor (tabla 5). De hecho, la descripción de la morfología del tercio facial medio de H. erectus clásico de Yunxian (Li y Etler, 1992) es la misma que la descrita para ATD6-69 (Bermúdez de Castro et al. 1997): “[…] el rostro es plano y ortognático, con prognatismo alveolar moderado; el maxilar tiene una fosa canina distintiva; la cara lateral del maxilar está orientada coronalmente y forma un gran ángulo con el zigomático; la raíz zigomática tiene un origen elevado; presenta un borde zigomaticomaxilar con una incisura malaris pronunciada […]” (Li y Etler, 1992, p. 404). La misma morfología es la que se encuentra en el rostro de Tangshan (Wang y Tobias, 2000 a,b, 2001; Liu et al., 2005). Además, en la tabla 5 se puede observar que los resultados de la morfología del tercio facial medio en H. erectus, sensu lato son muy variables, pero, si dividimos este grupo en H. erectus primitivos (u H. ergaster) y H. erectus clásicos, se aprecia que la morfología del primer grupo es más primitiva, mientras que la del segundo grupo es más derivada, lo que justifica la división de H. erectus, sensu lato, en estos dos grupos. Por lo tanto, la presencia de un tercio facial medio flexionado es una característica plesiomórfica de todos los hominoideos africanos actuales (Gorilla y Pan), australopitecos del este africano y primeros Homo (excepto para el Grupo 1470). Es por ello que estamos de acuerdo con Wang y Tobias (2000a), quienes afirmaron que el rostro flexionado es plesiomórfico y, por lo tanto, no se considera una base taxonómica efectiva para la nueva especie propuesta, Homo antecessor.


Figura 5. Morfología del tercio facial medio en homininos comparada con la de ATD6-69. En todas las imágenes, la línea roja marca la inclinación del plano infraorbital. A) Primeros Homo, Grupo 1470. Fila superior (vista frontal), izquierda: KNM-ER 1470, derecha: ATD6-69. Obsérvese en ER 1470 la ausencia de fosa canina, la práctica ausencia de flexión maxilar, y una cresta zigomaticoalveolar rectilínea oblicuamente orientada. Fila inferior (vista lateral), izquierda: KNM-ER 1470, derecha: ATD6-69; el plano infraorbital en ER 1470 se orienta anteroinferiormente (uniplanar), lo que indica un plano coronal comprimido con una cresta zigomaticoalveolar que nace desde abajo y hacia adelante. B) Primeros Homo, Grupo 1813. Fila superior (vista frontal), izquierda: KNM-ER 1813, derecha: ATD6-69. Obsérvese en ER 1813 la presencia de una fosa canina poco profunda y extendida, la baja flexión maxilar, y una cresta zigomaticoalveolar arqueada, sin incisura malaris. Fila inferior (vista lateral), izquierda: KNM-ER 1813 (imagen flip), derecha: ATD6-69; el plano infraorbital en ER 1813 se orienta posteroinferiormente, con un nacimiento ligeramente adelantado de la cresta zigomáticoalveolar. C) Homo erectus primitivos (u Homo ergaster). Fila superior (vista frontal), izquierda: KNM-WT 15000, derecha: ATD6-69. Obsérvese en WT 15000 la ausencia de fosa canina y la presencia de surco maxilar, y una cresta zigomaticoalveolar rectilínea orientada oblicuamente. Fila inferior (vista lateral), izquierda: KNM-WT 15000, derecha: ATD6-69; el plano infraorbital en WT 15000 se orienta anteroinferiormente (uniplanar), lo que indica un plano coronal comprimido con un nacimiento ligeramente retrasado de la cresta zigomaticoalveolar. D) Homo erectus clásico. Fila superior (vista frontal), izquierda: cráneo de Tangshan (Nanjing), derecha: ATD6-69. Obsérvese en Tangshan la presencia de una marcada fosa canina, la acusada flexión maxilar, y una cresta zigomaticoalveolar arqueada con incisura malaris. Fila inferior (vista lateral), izquierda: reconstrucción del cráneo de Zhoukoudian (Sawyer y Tattersall, 1995; Tattersall y Sawyer, 1996), derecha: ATD6-69; el plano infraorbital en Zhoukoudian se orienta posteroinferiormente, con un nacimiento muy retrasado de la cresta zigomaticoalveolar. Como puede apreciarse, la morfología de H. erectus clásico es idéntica a la de ATD6-69. Fotografías de ATD6-69 por cortesía de Roberto Sáez
En cuanto a la entrada nasal, ATD6-69 presenta la categoría 3 de Gower (Franciscus 2003), el modelo más abundante en humanos modernos (el 35,93% de una serie de 109 cráneos modernos [Franciscus, 2003]). Sin embargo, en humanos modernos, todos los modelos de combinaciones de crestas para formar el margen nasal inferior están presentes. Es más, la categoría 3 es ancestral en Homo, ya que se presenta en OH 62, KNM ER 3733 y KNM-WT 15000 (Franciscus, 2003). Por otra parte, algunos investigadores han demostrado que la morfología nasal de H. erectus primitivo (u H. ergaster) es prácticamente idéntica a la de los humanos modernos (Franciscus y Trinkaus, 1988), basándose en las siguientes características:
- presencia de una prominente espina nasal anterior en ciertos especímenes (Sangiran 4 [Franciscus y Trinkaus, 1988; Rightmire, 1998b; McCollum, 2000; Schwartz y Tattersall, 2003]; el cráneo de Gongwangling [Woo, 1966; Rightmire, 1998b])
- entrada plana de la cavidad nasal con alféizar nasal horizontal
- espina posterior
- en Sangiran 4 (el único espécimen en el que podemos observarla), se da también una inserción vomeral por encima del alféizar nasal.
Otra característica importante de la morfología del tercio facial medio de H. antecessor es la presencia de un tubérculo zigomaxilar. Arsuaga et al. (1999), Carbonell et al. (2005) y Bermúdez de Castro et al. (2017) apuntan que esta es una característica derivada y que se observa por primera vez en los restos de Gran Dolina y Zhoukoudian. Sin embargo, la presencia del tubérculo zigomaxilar ya había sido constatada por vez primera en KNM-ER 3733 —Pope 1991; Leakey y Walker 1985— (datado en ~1,8 Ma [Wood, 1991]) y Sangiran 17 —Pope 1991; Rightmire 1998b— (datado entre 1,25 y 1,30 Ma [Larick et al., 2001, 2004; Antón et al., 2007]). De esta manera, la aparición del tubérculo es anterior a su presencia en ATD6 y en los homininos de Zhoukoudian. Por lo tanto, la presencia de tubérculo zigomaxilar en H. antecessor es también un rasgo plesiomórfico, no derivado.
La última característica del tercio facial medio descrito como derivado en ATD6-69 es la presencia de un patrón de remodelado óseo de tipo moderno (Lacruz et al., 2013; Bermúdez de Castro et al., 2017). El patrón de remodelado óseo del tercio facial medio observado en ATD6-69 está presente en H. sapiens. En ambos, ATD6-69 y H. sapiens, se asocia con un rostro ortognático, una inflexión maxilar totalmente flexionada (que proyecta los huesos de la nariz hacia adelante), una región infraorbital orientada coronalmente con fosa canina, y una cresta zigomaticoalveolar curvada con incisura malaris (Lacruz et al., 2013). El rostro ortognático se asocia con áreas de reabsorción durante la ontogenia (Enlow y Wang, 1965; Enlow, 1966). En ATD6-69, estas áreas de reabsorción se localizan en el clivus nasomaxilar, la región anterolateral del maxilar y la fosa canina (fig. 2). Las áreas se deposición en ATD6-69 están próximas a la prominencia canina, en las paredes laterales de la abertura nasal y en la sección anterior del zigomático (Lacruz et al., 2013) (fig. 2).
El patrón de remodelado óseo facial en ATD6-69 y en H. sapiens es muy diferente del presente en los australopitecos (LH 2, LH 21, AL 333-105, Sts 2, Stw 59, Taung, Sts 24, Sts 57, MLD 2 y Sts 52), primeros Homo (OH 13, SK 27) y H. erectus primitivos (u H. ergaster) (KNM-WT 15000). En todos estos últimos especímenes observamos una gran zona de deposición denle el maxilar y en el clivus nasoalveolar, que contribuye al desarrollo de un perfil prognático (Bromage, 1989; Lacruz et al., 2015). Sin embargo, el patrón de remodelado óseo facial en Australopithecus sediba (MH1), que presenta un moderado prognatismo, es idéntico al de H. sapiens, que muestra bandas verticales de reabsorción de hueso en su cara inferior (Lacruz et al., 2015). De acuerdo con los autores, estas bandas verticales de reabsorción de hueso no son más que reminiscencias de las franjas alternantes de alta y baja tensión observadas en las raíces de los dientes anteriores durante la mordida, al menos tal y como son simulados en un modelo craneal humano sujeto al análisis de elementos finitos (Wang et al., 2010; Lacruz et al., 2015). Es más, la variabilidad de esta característica debería ser tenida en cuenta. En el estudio de Kurihara et al. (1980), se consideró una muestra de 27 cráneos de humanos modernos de entre 2 y 14 años de edad, de los que 20 (74%) muestran una cara anteroinferior con reabsorción predominante; en los restantes 7 especímenes (26%), se encontró que la mayor parte de la cara inferior era depositoria. Además, en una muestra de 33 chimpancés subadultos y crías, McCollum (2008) descubrió un patrón completo de deposición en la cara anteroinferior en solo 6 especímenes, mientras que en 21 de ellos el patrón era parcialmente reabsortivo, y completamente reabsortivo en cuatro. El mismo autor, en una muestra de 22 cráneos humanos modernos, muestra que en el 55% “se descubrió que mostraban grandes zonas de deposición superficial a lo largo del clivus nasoalveolar y el maxilar anterior” (McCollum 2008). Estos estudios nos indican que existe una estrecha relación entre un rostro ortognático y las áreas de reabsorción de hueso durante la ontogenia, localizadas en las zonas del clivus nasomaxilar, maxilar anterolateral y en la fosa canina. Y a la inversa, la deposición de hueso localizada en estas mismas regiones faciales no siempre se hace patente. De este modo, se produce una variabilidad en los modelos de reabsorción y deposición en el clivus nasomaxilar, maxilar anterolateral y en la fosa canina, que pueden presentarse tanto en rostros ortognáticos como en rostros prognáticos.
Aunque no ha sido posible establecer una serie ontogénica para H. erectus clásico basada en formas adultas del tercio facial medio, es razonable argumentar que los procesos ontogénicos de dicho tercio facial medio deben ser similares a aquellos encontrados en algunos humanos modernos (Wang, 1998). Por lo tanto, la presencia de un patrón de remodelado óseo facial en Paranthropus, Australopithecus sediba, ATD6-69 y H. sapiens podría indicar la aparición de este modelo en diferentes momentos de la evolución humana. En relación con la morfología del hueso temporal, según Arsuaga et al. (1999), la condición ancestral sería una escama temporal baja y con un borde superior recto. De acuerdo con estos autores, tal morfología está presente en H. habilis, H. erectus primitivos (u H. ergaster) y H. erectus clásico; mientras que en H. antecesor, la presencia de una escama temporal con un borde superior arqueado representa la condición derivada, que también se observa en los homininos del Pleistoceno medio europeo y africano, en los del Pleistoceno medio asiático, en neardentales y en humanos modernos. A este respecto, Carbonell et al. (2005) añaden que H. antecessor y H. erectus clásico tuvieron que compartir un hipotético ancestro común (posiblemente, H. erectus primitivo [u H. ergaster]) que conservara la primitiva condición de una escama temporal pequeña y plana, y apuntan que estas características se conservaron en H. erectus clásico.
Sin embargo, el cráneo de H. erectus clásico de Bouri (~1 Ma) ya presenta una escama temporal elevada y arqueada (información suplementaria en Asfaw et al., 2002). Igualmente, muchos ejemplares de H. erectus clásico, como los de Yunxian 2, Localidad 1 de Zhoukoudian (cráneos III, V, XI, y XII), y Tangshan (Wu y Poirier, 1995; Rightmire, 1998a; Liu et al., 2005; Etler, 2004), presentan también la misma morfología. Así, una escama temporal con un borde superior convexo ya ha sido encontrada en H. erectus clásicos de África y China. Por lo tanto, en lo que a la morfología craneal y del tercio facial medio respecta, los homininos ATD6 y H. erectus clásico comparten las características siguientes: presencia de fosa canina, cresta zigomaticoalveolar arqueada con incisura malaris y una raíz zigomática elevada, inflexión maxilar y morfología nasal prácticamente idéntica a la de los humanos modernos; así como un borde superior convexo de la escama temporal. Es por ello que la topografía de ATD6-69, considerada como derivada (Bermúdez de Castro et al., 1997; 2017; Arsuaga et al., 1999) es realmente un rasgo plesiomórfico, porque ya está presente en H. erectus clásico penecontemporáneo. Más aún, en un estudio de Freidline et al. (2013) sobre la evaluación de los cambios en la forma de desarrollo facial de H. antecessor, estos autores llegan a la conclusión de que la mayor parte de las características faciales similares a las de los humanos actuales presentes en ATD6-69, están interconectadas y pueden ser consideradas un vestigio ancestral en relación con humanos posteriores. Sugieren, además, que tales características ya estaban presentes en diferentes grados en más formas plesiomórficas, caso de H. erectus clásico y H. antecessor, y se han conservado en los humanos modernos.
CONCLUSIONES
Nuestra conclusión es que ninguna de las características del tercio facial medio que han sido utilizadas para definir a H. antecessor son exclusivas de los homininos de Gran Dolina. Todas de ellas se han hallado en H. erectus clásico. Por lo tanto, resulta insostenible separar ambos grupos en dos taxones diferentes, H. antecessor y H. erectus clásico; así, todos los homininos ATD6 de Atapuerca deberían ser clasificados como H. erectus clásico (Wang, 1998; Wang y Tobias, 2000a; Etler, 2004; Rabadà, 2005; Ribot et al., 2006, 2007, 2018; Ribot y García Bartual, 2016). Por lo tanto, los homininos ATD6 demonstrarían la presencia de H. erectus clásico en el Pleistoceno inferior superior y medio en Europa (Wang y Tobias, 2000a; Rabadà, 2005; Ribot y García Bartual, 2016; Ribot et al., 2018). Es más, el rostro bajo y flexionado con un prognatismo ligero o ausente y que es propio de los humanos modernos pudo desarrollarse fácilmente a partir de un rostro bajo y flexionado como el que presentaba H. erectus clásico.
AGRADECIMIENTOS
Queremos agradecer a los Doctores Vance Hutchinson, Esteban Sarmiento, Bill Jungers, Carles Ferrández, Lluís Gibert, Jordi Galbany y al Profesor Xinzhi Wu sus revisiones del manuscrito y sus útiles sugerencias.
Al Dr. Ian Tattersall por habernos proporcionado información sobre el maxilar de Homo erectus de Zhoukoudian, al Dr. Scott Maddux por su información sobre la variabilidad de la cresta zigomaticoalveolar, y al Dr. Jeffrey Schwartz por la información facilitada sobre el maxilar de KNM-ER 1813.
Damos las gracias al Dr. Alejandro Pérez-Pérez por habernos permitido estudiar los moldes de homininos del Departamento de Biología de la Evolución, Ecología y Ciencias Ambientales (Sección de Zoología y Antropología Biológica) de la Universidad de Barcelona, y al Dr. Domènec Campillo por hacer lo propio con los moldes de homininos y colecciones de cráneos humanos actuales del Laboratorio de Paleopatología y Paleoantropología (Museo Nacional de Arqueología de Cataluña, en Barcelona).
Imágenes de ATD6-69 de las figuras 1 y 5 por cortesía de Roberto Sáez. Imágenes del Homo erectus de Zhoukoudian por cortesía del Dr. Ian Tattersall y de Gary Sawyer.
Queremos dar las gracias, también, a los dos revisores anónimos por sus comentarios y sugerencias.
Contribuciones de los autores
Como investigador principal, F.R.T. ha diseñado y desarrollado el estudio, y concebido y diseñado el manuscrito; M.G.B. ha puesto en orden y ha analizado los datos, y escrito parte del manuscrito; Q.W. ha escrito parte del manuscrito, y lo ha corregido. Todos los autores han leído y aprobado el manuscrito final.
Conflicto de intereses
Los autores declaran que no existe conflicto alguno de intereses.
Artículo Original en Content Sciendo


GLOSARIO:
- Hipodigma: Todo el material fósil que se conoce de una especie
- Filogenético: Perteneciente o relativo a la filogenia.
- Filogenia: Del Griego φῦλον phŷlon ‘raza’ y -genia. Parte de la Biología que se ocupa de las relaciones de parentesco entre los distintos grupos de seres vivos. Origen y desarrollo evolutivo de las especies, y en general, de las estirpes de seres vivos.
- Taxón: Del Griego τάξις, “táxis”, ordenamiento, es un grupo de organismos emparentados que en una clasificación dada han sido agrupados, asignándole al grupo un nombre en latín, una descripción si es una especie, y un tipo. Cada descripción formal de un taxón es asociada al nombre del autor o autores que la realizan, los cuales se hacen figurar detrás del nombre. En latín el plural de taxón es taxa, y es como suele usarse en inglés, pero en español el plural adecuado es taxones o táxones». La disciplina que define a los taxones se llama Taxonomía.
- Fosa canina: Definida por Mellinger (1940), es la depresión infraorbital que incluye la mayor parte de los procesos cigomáticos de la mandíbula
- Cigomaxilar: Relativo al hueso cigomático
- Hueso cigomático: También denominado “malar”, es un hueso par, corto y compacto situado en la parte más externa de la cara. Forma el pómulo de la cara y parte de la órbita y presenta un saliente o proceso cigomático que se une hacia atrás con el proceso cigomático del hueso temporal. Solo se denomina así en los mamíferos y se corresponde con el hueso yugal del resto de los vertebrados. Se articula con el temporal, el maxilar, el esfenoides y el frontal.
- Apomórfico: De apomorfia o apomorfía
- Apomorfia: Rasgo o carácter biológico evolutivamente novedoso. Una novedad evolutiva derivada de otro rasgo perteneciente a un taxón ancestral filogenéticamente próximo. El concepto de apomorfia se opone al de plesiomorfia, que se refiere a los rasgos históricamente más antiguos, de los cuales derivan las apomorfias
- Subalveolar: Bajo un alvéolo
- Alvéolo: Cavidad dentro del hueso maxilar en la que está encajada el diente
- Zhoukoudian: Conjunto de cuevas en el distrito suburbano de Fangshan, en Beijing. Protagonista de muchos descubrimientos arqueológicos, incluyendo uno de los primeros especímenes de Homo erectus (Homo erectus pekinensis), el Hombre de Pekín, así como un surtido de huesos de la hiena gigante Pachycrocuta brevirostris
- Ortognático: Del Griego, “orthos“, recto, y “gnathos“, mandíbula. Así, la cirugía ortognática tendrá como finalidad mover la mandíbula a una posición más adecuada, más estética, más funcional y por supuesto más saludable; ésto se logra a través de la combinación de dos áreas o especialidades médico-odontológicas: la ortodoncia mueve los dientes y la cirugía maxilofacial mueve los huesos. Las deformidades dentofaciales son muy frecuentes, entre un 8 y un 12 por ciento de la población mundial presenta algún tipo de desequilibrio relacionado con los dientes y el rostro, es decir, padecen de disarmonías dentofaciales, alteraciones que afectan tanto a los dientes como a la cara
- Rasgos derivados: Son rasgos evolucionados respecto al inmediato antecesor
- Rasgos plesiomórficos: Frente a los anteriores, son rasgos primitivos
- Clivus nasoalveolar o subnasal: Porción del cráneo localizada sagitalmente que se extiende desde la unión anterior del tabique nasal hasta el margen de las premaxilas en el punto alveolar osteométrico
- Deposición: Acumulación de material, en este caso, óseo
- Tercio facial medio: Ver imagen
- Tubérculo cigomaxilar: Ver imagen
- Patrones de surco no Y: Patrón que se presenta en los molares inferiores en forma de Y
- Trigónido medio: Referido a la morfología de los molares inferiores
- Fóvea: Estructuras presentes en los dientes, concretamente, en premolares, tal y como el artículo refiere
- Incisura malar: Muesca o fuerte incurvación en la cresta cigomaticoalveolar
- Línea bicanina: Línea imaginaria entre los caninos superiores que permite observar la ubicación de los incisivos, si están muy adelantados o poco
- Escama: Se refiere a la escama del hueso temporal. Este hueso está dividido en dos porciones, la porción escamosa y la porción petrosa
- Hueso temporal: Ver imagen
- Sinapomorfia: Una sinapomorfia o sinapomorfía es un carácter homólogo apomórfico compartido por todos los individuos de un taxón. Es decir, una sinapomorfía es una novedad evolutiva que permite diferenciar a un taxón de otros taxones
- Penecontemporáneo: Casi contemporáneo
- Patrón del surco cortical: Surco de la corteza cerebral para alojar venas
- Análisis cladístico: Que parte de los principios de la Cladística
- Cladística: Del Griego κλάδος, klados, rama, es la especialidad de la Biología que define las relaciones evolutivas entre los organismos basándose en similitudes derivadas [Ver: Rasgos derivados]. Es la más importante de las sistemáticas filogenéticas que estudian las relaciones evolutivas entre los organismos
- Dmanisi: Yacimiento arqueológico situado en la república de Georgia, al sur de Tiflis, en Próximo Oriente, en el que se encuentra una larga secuencia estratigráfica que va desde el Pleistoceno inferior hasta la Edad Media. De hecho, en los niveles más antiguos, datados en no menos de 1’8 millones de años, se ha exhumado tanto restos de industria lítica, como abundantes fósiles de animales y lo que podrían ser los restos más antiguos de seres humanos hallados fuera de África
- Sangiran: Yacimiento arqueológico ubicado en Java, en Indonesia. Según un informe de la UNESCO de 1995, “Sangiran ha sido reconocido por la comunidad científica como uno de los más importantes del mundo en cuanto al estudio de fósiles humanos, a la altura de Zhoukoudian (China), Willandra Lakes (Australia), Olduvai Gorge (Tanzania) y Sterkfontein (South Africa), y más fructífero en hallazgos que todos los anteriores
- Gongwangling: Yacimiento del condado de Lantian, en China, donde se encontró a una subespecie de Homo erectus, Homo erectus lantianensis, el llamado Hombre de Lantian. Descubierto en 1963, fue descrito por primera vez por J. K. Woo al año siguiente. Su capacidad craneal era de 780 centímetros cúbicos, similar a la de su contemporáneo, el Hombre de Java
- Yuxian: También conocido como el Condado de Yu, Yuxian se encuentra al este de la provincia china de Shanxi
- Mediofacial: Del tercio facial medio
- Cigión: Punto antropométrico que se encuentra muy cerca de la sutura cigomaticomaxilar, y conforma la parte lateral más externa del arco cigomático, de la cara
- Sagital: Plano sagital, que divide el cuerpo perpendicularmente en dos mitades exactamente iguales
- Evertido: Una estructura evertida es aquella que sobresale del plano en que se encuentra al girar sobre el mismo
- Morfotipos: Tipos de morfología en un mismo género. Dos morfotipos indican dos posibles especies
- Proceso cigomático: En Osteología, una apófisis o proceso cigomático es una parte que “surge de”
- Piriforme: En forma de pera
- Hominoides: Se corresponden con la superfamilia Hominoidea. Incluye a los homínidos y a los hilobátidos (familia Hylobatidae, de la que sólo viven los gibones)
- Tuberosidad: Zona donde hueso y tendón se unen
- Cresta turbinal: Cresta que se aloja en la apertura nasal y que se origina cerca del extremo anterior del cornete inferior
- Ontogenia: Desarrollo del individuo, en especial, el que sucede durante el período embrionario
- Rostro prognático: Que presenta prognatismo
- Prognatismo: Del Griego πρo y γναθος, mandíbula adelante, consiste en una deformación de la mandíbula por la cual ésta está adelantada respecto a su posición normal
- Corrugación: Arruga
- P3/ P4/ M1: P3, primer premolar superior; P4, segundo premolar superior; M1, primer molar superior
- Jugum: Juga o eminencia de los dientes, especialmente, de los superiores. La eminencia canina es un “tubo” por el que se despliega la raíz del canino y que asciende junto al margen lateral nasal
- Foramen infraorbital: Plano infraorbital
- Cara flexionada: Propia de los humanos modernos, frente a la cara plana o lisa, aquella que no presenta flexiones – por ejemplo, la de Homo rudolfensis o neanderthalensis
- Medialmente: Que está situado cerca de o en el plano medio
- Elongado: Estirado
- Masetero: Músculo implicado en la masticación, que se extiende desde el arco cigomático anterior y desciende por toda la rama de la mandíbula
- Epigenético: Relativo a la Epigenética
- Epigenética: Estudio de modificaciones en la expresión de genes que no obedecen a una alteración de la secuencia del ADN y que son heredables. Fuentes de modificaciones importantes de los genes son los factores ambientales, que pueden afectar a uno o varios genes con múltiples funciones.
- Análisis de elementos finitos: Análisis empleado en Biomecánica para conocer cómo se distribuyen las fuerzas de estrés por los huesos, en este caso, faciales
- Sensu lato: En sentido más amplio
- Uniplanar: Se trata de un plano que se dispone en línea recta, es decir, su parte inferior ni se desplaza hacia adelante ni hacia atrás
- Ancestral: Primitivo
- Vomeral: Hueso de la nariz. Ver imagen
- Reabsortivo: Relativo a un proceso de reabsorción


Galería de imágenes y webgrafía:
- El Glosario ha sido elaborado, básicamente, con las aportaciones del Dr. Ribot. Además, con las consultas que hemos realizado en páginas como Real Academia Española, Wikipedia en Español y English Wikipedia
- Otras páginas consultadas han sido Diario de Atapuerca y Terra Antiqvae
- Las fotografías de D. Roberto Sáez las hemos tomado de Nutcrackerman
- Imágenes restantes tomadas de DocPlayer.es y Slideplayer.es
- El ‘Homo antecessor’, descubierto en Atapuerca, se extinguió sin descendencia
Estimados lectores:
Hemos corregido el error que afectaba al enlace que debía redirigirnos a la publicación original. Muchas gracias por vuestro apoyo.
El equipo de LPDLA