Cuentan las crónicas que esta fue la arenga con la que el General y estadista cartaginés Aníbal Barca (247 – 183 a.C.) lanzó a sus tropas a la Segunda Guerra Púnica contra Roma en 218 a.C. Las mismas que un año antes le habían dado la victoria contra Sagunto, permitiéndole el control de la costa levantina.
Perfectamente pertrechados con manteletes – grandes escudos de madera ensamblados como un rompecabezas y de formas diversas, cuadrangulares, circulares -, ballestas, y picos, los soldados cartagineses, con Aníbal al frente, derribaron las murallas de la ciudad mediante el empleo de catapultas, parapetos móviles y las torres de asalto que los griegos llamaban helepolis, las tomadoras de ciudades.
Se iniciaba, así, un nuevo conflicto bélico entre Cartago y Roma, que Aníbal prepararía desde su residencia en Qart Hadasht encomendándose al dios Melqart.
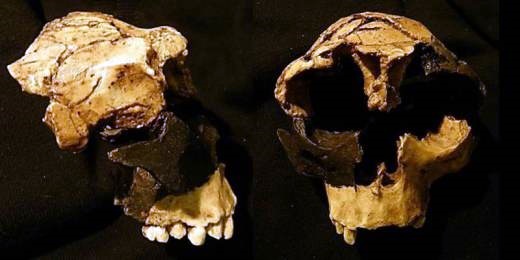
Homo antecessor. Composición de ATD6-15 y ATD6-69. Fotografía de Roberto Sáez
Hace casi veinticinco años que el equipo de Atapuerca, liderado por los paleoantropólogos Juan Luis Arsuaga, Jose María Bermúdez de Castro y Eudald Carbonell, anunciaba oficialmente el descubrimiento de una nueva especie en el genero Homo a la que denominaron ‘antecessor’, ‘pionero’, término que Bermúdez de Castro escogió al rememorar las avanzadillas que el Imperio Romano enviaba a tierras ignotas. La noticia revolucionó a la comunidad científica y puso al yacimiento burgalés en la primera línea de la investigación paleoantropológica mundial. Sin embargo, la juventud del espécimen ATD6-69, base de sus investigaciones, pronto llevaría a otros colegas de profesión a plantearse serias dudas acerca de la viabilidad de antecessor como nueva especie. Y surgió la pregunta, ¿existió realmente Homo antecessor?
Para empezar esta nueva temporada de publicaciones, os traemos hoy un trabajo muy interesante que firma nuestro amigo Daniel Salmador Hernández.
Daniel es miembro del grupo Keltika Hispanna, cuyas canciones han formado parte, en alguna ocasión, de Ecos de Paisajes Sonoros, la lista de reproducción del blog. Apasionado por el Folclore y la Etnografía españolas, es uno de los fundadores de la asociación Raíz Ibérica, cuyas actividades están centradas en la recuperación, divulgación y preservación del rico patrimonio cultural de la Península Ibérica.
En “Las Moras y Moros Mitológicos”, Daniel se vale de la Toponimia para hacer un seguimiento de la presencia de estos personajes de leyenda a lo largo y ancho de la Piel de Toro, al tiempo que contrasta todas esas historias con tradiciones culturales posteriores. Los moros de los que aquí se habla nada tienen que ver con los habitantes del Magreb, como muchos podrían pensar a primera vista. Hemos de decir, también, que el término “moro” no tiene en sí mismo una acepción peyorativa, pues “moro” deriva del latín “mauri”, palabra que designaba a los habitantes de la Mauritana Tingitana, la provincia romana constituida en el Norte de África, tal y como Estrabón relata.
Por último, y a fin de facilitar su lectura, hemos tirado de diccionario para elaborar un glosario de términos que sigue su orden de aparición en el artículo. Esperando que éste sea de vuestro agrado, os dejamos ya con“Las Moras y los Moros Mitológicos”.
Glosario de términos:
Hilandera: Persona que tiene por oficio hilar. Lugar donde se hila
Sincretismo: Combinación de distintas teorías, actitudes u opiniones. Sistema filosófico que trata de conciliar doctrinas diferentes. Expresión en una sola forma de dos o más elementos lingüísticos diferentes
Cuélebre: Dragón fabuloso de la mitología asturiana
Castro: Poblado fortificado celta, por lo general prerromano, aunque existen ejemplos posteriores que perduraron hasta la Edad Media en Europa. Propios de finales de la Edad del Bronce y de la Edad del Hierro, son muy comunes en la Península Ibérica, sobre todo en el noroeste, con la cultura castreña, y en la Meseta, con la cultura de las cogotas. El término castro proviene del latín “castrum”, que significa “fortificación militar”. LLamado también “oppidum” (en plural, “oppida”) cuando son de gran tamaño
Megalito: Monumento prehistórico construido con grandes piedras sin labrar
Petroglifo: Figura hecha por incisión en roca realizada por pueblos prehistóricos
Libación: Acción de derramar vino u otro licor en honor de alguna divinidad. Entre los antiguos paganos, ceremonia religiosa que consistía en derramar vino u otro licor en honor de los dioses
Rueca: Instrumento que sirve para hilar y que se compone de una vara delgada con un rocadero hacia la extremidad superior
Fusa: Ver “huso”
Soliforme: Manifestación de Arte Prehistórico Esquemático que representa al Sol resplandeciente
Dolmen: Monumento megalítico compuesto de una o más lajas colocadas de plano sobre dos o más piedras verticales
Talayot: Monumento megalítico de las Islas Baleares semejante a una torre de poca altura
Taula: Monumento megalítico frecuente en Menorca, constituido por una piedra colocada verticalmente en el suelo que soporta otra plana horizontal, a modo de mesa
Menhir: Monumento megalítico que consiste en una piedra alargada colocada verticalmente en el suelo
Espelunca: Cueva, gruta, concavidad tenebrosa
Baladre: Adelfa
Tragantía: Leyenda del pueblo de Cazorla, en Jaén, sobre la reconquista de la ciudad y que cuenta la historia de una princesa mora encerrada en una cueva próxima al castillo por su padre, el rey, ante avance inexorable de las tropas cristianas. Ordenó tapiar la entrada de la cueva con la intención de rescatarla cuando las tropas cristianas ya se hubieran marchado, pero, éstas alcanzaron al rey y a su séquito, les dieron muerte a todos y ocuparon la población de Cazorla. Según cuenta la leyenda, la princesa, aprisionada en la cavidad, ignorada por todos, según cuenta la leyenda, metamorfoseó en mitad lagarto, mitad mujer a causa de la oscuridad y de la humedad y, desde entonces, permanece en la cueva de la que sólo sale cada noche de San Juan, en la que se puede oír su lamento: “Yo soy la tragantía, hija del rey moro, quien me oiga cantar no verá la luz del día ni la noche de San Juan”
Semita: Según la tradición bíblica, descendiente de Sem, uno de los hijos de Noé. Que pertenece a alguno de los pueblos que integran la familia formada por los árabes, los hebreos y otros pueblos
Arquetipo: Modelo original y primario en un arte u otra materia. En Psicología, representación que se considera modelo de cualquier manifestación de la realidad; imagen esquema congénito con valor simbólico que forma parte del inconsciente colectivo. En Religión, tipo soberano y eterno que sirve de ejemplar y modelo al entendimiento y a la voluntad humanos.
Huso: Instrumento manual, generalmente de madera, de forma redondeada, más largo que grueso, que va adelgazándose desde el medio hacia las dos puntas y sirve para hilar torciendo la hebra y devanando en él lo hilado. También, instrumento, algo más grueso y más largo que el huso de hilar, que sirve para unir y retorcer dos o más hilos. Además, instrumento de hierro, como de medio metro de largo y del grueso de un bellote, que tiene en la parte inferior una cabezuela, también de hierro, para que haga contrapeso a la mano, y sirve para devanar la seda. En Heráldica, losange largo y estrecho. En Tecnología, cilindro de un torno
Tríada: Conjunto de tres seres o de tres objetos estrecha o especialmente vinculados entre sí
Homofonía: En Lingüística, cualidad de homófono. En Música, conjunto de voces que cantan al unísono, o de sonidos simultáneos.
Toponimia: Conjunto de los nombres propios de un lugar, país o región. Rama de la Onomástica que estudia el origen de los nombres propios de lugar, así como el significado de sus étimos
Ciclópeo: Perteneciente o relativo a los cíclopes. Dicho de ciertas construcciones antiquísimas que se distinguen por el enorme tamaño de sus piedras, unidas por lo común sin argamasa. Gigantesco. Excesivo o muy sobresaliente
Etnológico: Perteneciente o relativo a la Etnología, la ciencia que estudia comparativamente los orígenes y expresiones de la cultura de los pueblos, a partir de los datos proporcionados por la Etnografía. Ésta estudia, de manera descriptiva, la cultura popular.
Feérico: Perteneciente o relativo a las hadas
Referencias bibliográficas consultadas para el glosario:
“… Ha caído la noche y los bárbaros no han venido… Y entonces ahora, ¿qué va a pasar con nosotros sin los bárbaros? Al menos esa gente era una cierta solución”
Cavafis, “Esperando a los Bárbaros”
“Bárbaros y Romanos en Hispania [400 – 507 A.D.]” es una obra del Profesor Javier Arce Martínez publicada por vez primera en el año 2015 por la editorial Marcial Pons Historia. A lo largo de sus más de trescientas páginas, el autor hace un exhaustivo recorrido por un siglo tan determinante para la Historia de España como lo fue el siglo V d.C.
Javier Arce Martínez (1945) es un historiador y arqueólogo español muy reconocido en el entorno académico, tanto nacional como internacional. Ejerce su tarea docente e investigadora en instituciones tan prestigiosas como el Consejo Superior de Investigaciones Científicas (CSIC), donde trabaja como Profesor de Investigación, habiendo dirigido la Escuela Española de Historia y Arqueología a él adscrita en Roma, y la Universidad de Lille, en Francia, donde imparte sus enseñanzas como Profesor de Arqueología Romana. Así mismo, ha coordinado junto con I. Wood y E. Chrysos el programa científico de la European Science Foundation “The Transformation of the Roman World”. Colaborador en numerosas publicaciones, es autor de una veintena de obras entre las que destacan “Funus Imperatorum: Los Funerales de los Emperadores Romanos” (1990), “El último Siglo de la Hispania Romana” (2009), “Esperando a los Árabes: Los Visigodos en Hispania (507-711)” (2013) o ésta que nos ocupa, motivo del presente trabajo, “Bárbaros y Romanos en Hispania 400-507 A.D” (2007). Sigue leyendo Bárbaros y Romanos en Hispania, 400 – 507 A.D. Recensión (I)
Como en épocas anteriores, el siglo V fue un periodo de inseguridad, sobre todo, en el entorno rural, donde ladrones y salteadores de caminos campaban a sus anchas. Así se refleja en las referencias que Consencio hace a los bárbaros que roban libros en su carta a Agustín de Hipona, y en los textos de Hidacio en los que menciona la presencia de piratas norteuropeos en el Cantábrico y trata la cuestión de la bacaudae, la bagauda. A lo que habría que sumar los episodios recurrentes de resistencia local frente al poder romano y la presencia goda.
Con motivo del descubrimiento de la Cueva de Altamira hace ya siglo y medio, en la localidad cántabra de Santilla del Mar, la plataforma Google Arts & Culture, en colaboración con el Museo Nacional y Centro de Investigación de Altamira, se ha descolgado con la publicación de una exposición digital y de un doodle para conmemorar tan importante aniversario.
Si bien sus descubridores, D. Marcelino Sanz de Sautuola y su hija María, ya la conocían desde 1868, no sería hasta 1875 cuando la niña accedió, por vez primera, a la sala que exhibe las magníficas representaciones que todos conocemos, en las que la pequeña creyó ver bueyes, y así se lo hizo saber a su padre.
Sin embargo, el hallazgo de la Capilla Sixtina del Arte Prehistórico, como ha sido denominada desde entonces, se tornó en un calvario para D. Marcelino, quien tuvo que soportar las críticas y el desprecio de una comunidad científica – francesa, especialmente – que, como en otras tantas ocasiones, en absoluto se hallaba preparada para aceptar una revelación tal. El valor de sus pinturas y el trabajo de D. Marcelino no serían reconocidos hasta muchos años después.
Padre e hija en unas fotografías de la época
Puesto que sólo un reducido número de visitantes al año puede visitar la cavidad original, a fin de evitar su rápido deterioro y garantizar su conservación, para facilitar el acceso masivo del público interesado en conocer esta joya de nuestro patrimonio cultural se construyó la llamada Neocueva, una réplica exacta de la primera. Inagurada en 2001, su contemplación resulta igualmente toda una experiencia, como tuvimos ocasión de comprobar en un viaje por tierras cántabras. Visita que recomendamos encarecidamente a todos aquellos que aún no la conozcan.
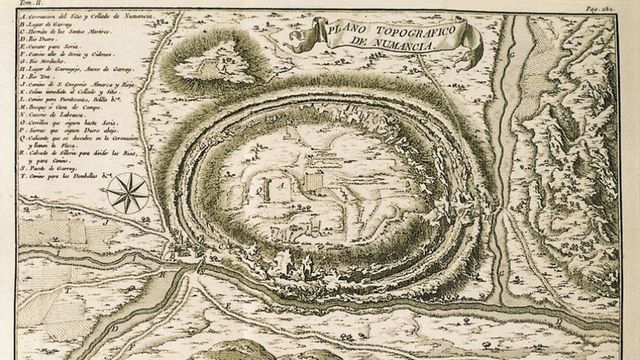
En este 2018, Año Europeo del Patrimonio Cultural, y habiendo celebrado en 2017 el 2150 Aniversario del Asedio y Caída de Numancia, se acaba de publicar un libro interactivo editado por Jose María Luzón y Mª del Carmen Alonso, con textos a cargo de Alfredo Jimeno Martínez, un trabajo de investigación histórica y arqueológica excelente en cuya confección se ha empleado las técnicas más modernas de prospección de yacimientos.
Representaciones de caballos en cerámicas numantinas, según Wattenberg, 1963
La obra se compone de ocho capítulos, que ponen a la ciudad en su contexto histórico, más un prólogo, un epílogo, y un apéndice. Este último nos ofrece toda información digital con la que se ha trabajado: Georradar, Fotografía 3D, Bases de Datos, así como Bibliografía y Hemeroteca. Con la posibilidad de ser leído directamente tanto en Inglés como en Alemán. Un proyecto, en nuestra opinión, ambicioso y de resultados más que meritorios, que podéis disfrutar en este enlace:
Hesperia. Del Latín Hesperĭus, y éste, del Griego ῾Εσπέριος, Hespérios.
Hesperia es uno de los nombres por los que se conoció a la Península Ibérica, más bien, a una parte de ella, antes que por el latinizado Hispania, al encontrarse aquélla al Oeste del mundo que los griegos conocían, en el Extremo Occidente.
Venus orbitando
Hesperia hace referencia al planeta Venus, que ellos llamaban Héspero o Véspero, que podía ser observado en dirección Oeste y durante la puesta de Sol si las condiciones meteorológicas eran propicias. Como Hesperia, bautizaron, también, a la Península Itálica.
Desplazamiento de Venus sobre la vertical de la Península Ibérica
Iberia, Hesperia, Hispania, términos todos que son prueba de la gran importancia, geoestratégica, sociocultural y comercial de la Península Ibérica en el Mundo Antiguo.
Íbero. Del Latín Ibērus, y éste del Griego Ιβηρ, Ιβηρος, Íbēr, Íbēros.
Íbero significa natural de Iberia, individuo perteneciente a alguno de los pueblos que se habían establecido en la Península Ibérica con anterioridad a la llegada de los primeros colonizadores griegos y fenicios. Pueblos que la ocuparon casi en su totalidad, desde la Bahía de Cádiz hasta el Mediodía de la Francia que hoy conocemos, con gran presencia en el Levante peninsular. La sociedad íbera se conformará durante un largo proceso que se inicia en el siglo VIII a.C. y que culminará en torno al año 500 a.C.
Recreación de la policromía de la Dama de Elche
Dada la imprecisión de la de Heródoto sobre la visita de Kolaiss de Samos a la Península Ibérica, la primera cita fiable es la de Avieno, quien empleará el término iberi para referirse a las tribus indígenas que dominaban la zona del Ebro (para los griegos, Iber, para los romanos, Hiberus), que otros autores como Pomponio Mela, Erastótenes o Estrabón citarán en sus escritos, también.
Los íberos o libioibéricos, como algunos especialistas los denominan, llegaron a ocupar zonas del Norte de África, y se constituyeron en el grupo racial más importante de la Península, lusitanos incluidos, presentando características propias en cada una de las zonas que dominaban. Las dos etnias originales, íberos y celtas, no dieron lugar al pueblo celtíbero, como reza la tradición. Era éste un pueblo autónomo, con su propia identidad, cuyos límites territorriales rebasaban ampliamente los del Ebro, tal y como los romanos constataron a su llegada a la Península a finales del siglo III a.C. y refieren los autores clásicos. Su influencia creciente, unida a la de los colonizadores grecofenicios, sentará las bases de la futura cultura hispánica.
Como lengua prerromana, el Íbero era el idioma que se hablaba en la Península hasta el siglo I d.C. Su adscripción resulta todavía desconocida. Por íbero entendemos, también, todo aquello relativo o perteneciente a la cultura del mismo nombre.
Para saber más de los íberos la bibliografía es muy abundante. Pero si nos permitís la recomendación, os proponemos la obra que en este enlace podéis descargar gratuitamente, titulada “Íberos: Sociedades y Territorios del Occidente Mediterráneo”, de Susana González Reyero, publicado por la prestigiosa editorial del Consejo Superior de Investigaciones Científicas, CSIC.
El Levantamiento del Dos de Mayo en Madrid o La Lucha con los Mamelucos
Dos de las mejores obras de Francisco de Goya y Lucientes, “El Levantamiento del Dos de Mayo de 1808“, y “Los Fusilamientos del Tres de Mayo de 1808“, comparten una misma temática, la de la insurrección de un pueblo, el español, subyugado por un monarca despótico, primeramente, por el invasor francés, más tarde, y las consecuencias más inmediatas que resultarían de todo ello.
Pintadas en 1814, en un intento, quizás, de congraciarse con Fernando VII, ambas obras ilustran el inicio y la deriva de la primera guerra fallida de todas en las que Napoleón Bonaparte se embarcaría por razón de sus ansias expansionistas.
Aquel Dos de Mayo, los habitantes de Madrid tomaron las calles de la ciudad. Pertrechados con estacas y cuchillos, se enfrentaron a las tropas invasoras, a su artillería y a la caballería mameluca de Joachim Murat, noble y militar francés, cuñado de Napoleón, bajo las que caerían masacrados en la misma Puerta del Sol. De madrugada, en las primeras horas del Día Tres, el pelotón de fusilamiento apostado en la montaña de Príncipe Pío completaría la matanza.
En “El Levantamiento del Dos de Mayo de 1808”, Goya procedió a localizar los acontecimientos con el máximo esmero. Es a los pies del edificio de Correos donde transcurre la acción, donde el espectador se convierte en “testigo ocular” de la matanza. En “Los Fusilamientos del Tres de Mayo de 1808”, el maestro pone de manifiesto, de manera eficaz, las contradicciones de la ideología de Bonaparte.
La exposición que hace de las ejecuciones ordenadas por Murat es la otra cara de la moneda de la iconografía davidiana, la brutal y triste realidad que el lustre imperial pretendería difuminar. Y es que, como si de un historiador de la perversidad se tratara, J.L. David dedicaría su obra a la exaltación y a la promoción del poder del Estado Revolucionario Francés. El empeño mostrado por David en poner su talento al servicio del Imperio y la glorificación de una figura, la de Napoleón, que había cernido la guerra sobre Europa, le granjearían no pocas críticas.
Los Fusilamientos del Tres de Mayo en Madrid
En “Los Fusilamientos del Tres de Mayo de 1808”, el pintor español logra un ejercicio de composición que resulta tan magistral como dramático. Vemos a los ejecutores dispuestos de perfil, de manera casi anónima, sin mostrar la fisonomía de sus rostros. Frente a ellos, los ajusticiados, que conforman tres grupos. Los que ya han sido fusilados, que yacen inermes sobre charcos de sangre. Los que están a punto de serlo, que representan el grupo más emotivo, realzado por el uso que el maestro Goya hace, sobre todo, de la luz y de la disposición de los personajes. Al fondo, una larga hilera de prisioneros, que espera su turno para enfrentarse indefectiblemente a la muerte, que allí mismo les aguarda.
Y es que la mayor muestra de la resistencia española se produjo a nivel local, por parte del clero y del campesinado. Si bien las Juntas intentarían organizar a los rebeldes, fueron las guerrillas las encargadas de hostigar a los franceses por toda la Península. Una guerra desigual, que acercaría al maestro a la figura de cronista, de reportero de guerra, como decimos ahora, más que a la de historiador. Una guerra que él retrataría, de manera exhaustiva, en la colección de aguafuertes denominada “Los Desastres de la Guerra“, compuesta por tres grupos principales.
Desastre Número 44, “Yo, lo vi”
Los dos primeros grupos pertenecen a época napoleónica, con la guerra y la hambruna como protagonistas. El tercero se recrea en el anticlericalismo de los “capuchos“, y pertenece al periodo de la Restauración reaccionaria.
La posición del testigo que contempla tales grabados ha dado lugar a dos interpretaciones contrapuestas. La primera de ellas se fundamenta en lo que Goya había escrito al pie del Desastre número 44, “Yo, lo vi” para asegurar que él mismo lo había visto, lo que le habría permitido elaborar todo un imaginario de la guerra.
La segunda interpretación sostiene que Goya no lo vio, otros instantes del conflicto, si, pero no los que estos grabados nos ofrecen. Para sus partidarios, son la creación personal desde unos acontecimientos que Don Francisco habría tomado como punto de partida. Un testigo, Goya, tan cerca de lo que realmente ocurrió como tan lejos, tan discutible como exento de toda sombra de duda.